We certify that we have read this
dissertation and that, in our opinion, it is satisfactory in scope and quality
as a dissertation for the degree of Doctor of Philosophy in Microbiology.
|
|
PHYSIOLOGICAL,
SEROLOGICAL, AND PLASMID CHARACTERIZATION
OF
FAST-GROWING RHIZOBIA THAT NODULATE SOYBEANS
A DISSERTATION SUBMITTED
TO THE GRADUATE DIVISION OF THE
UNIVERSITY OF HAWAII IN
PARTIAL FULFILLMENT
OF THE REQUIREMENTS FOR
THE DEGREE OF
DOCTOR
OF PHILOSOPHY
IN
MICROBIOLOGY
DECEMBER
1983
By
Michael
Jay Sadowsky
Dissertation
Committee:
B. Ben Bohlool, Chairman
Leslie R. Berger
Clair E. Folsome
John B. Hall
David M. Karl
We certify that we have read this
dissertation and that, in our opinion, it is satisfactory in scope and quality
as a dissertation for the degree of Doctor of Philosophy in Microbiology.
|
|
ACKNOWLEDGEMENTS
I am extremely grateful
to Dr. Ben Bohlool for his guidance, understanding, and constructive criticisms
throughout all phases of this research.
I would like to thank all
members of my dissertation committee for their helpful suggestions and
criticisms in the preparation of this dissertation.
Lastly, I would like to
thank fellow graduate students and friends Renee Kosslak, Mark Kingsley,
Stephen Dowdle, Paul Singleton, and Robert Woolfenden for their helpful
discussions, comments, friendship, and comradeship while working together in
Dr. Bohlool’s laboratory.
This research was funded
in part by grants SEA/AR-58-9AHZ-2-670 from the U.S. Department of Agriculture
and AID/DSAN-G-0100 from the U.S. Agency for International Development.
ABSTRACT
The newly described
fast-growing, acid-producing soybean rhizobia from The People’s Republic of
China (PRC) were examined to determine their degree of physiological and
serological relatedness to each other, to the "typical" slow-growing R.
japonicum, and to other fast-growing species of Rhizobium. The PRC strains were also investigated to
determine: 1) whether they contained high molecular weight plasmids; 2) if
there were structural relationships between plasmids from different strains; 3)
if plasmids from these strains are involved in the nodulation of soybeans; and
4) whether the fast-growing soybean strains were capable of accepting,
maintaining, and expressing symbiotic plasmids from other fast-growing species
of Rhizobium.
Results of these
investigations have indicated that: (1) While the fast-growing soybean rhizobia
share symbiotic host-specificity with the typical slow-growing R. japonicum
(they both nodulate the same host legume), they appear more closely related, on
a microbiological, biochemical, and physiological basis, to other fast-growing
species of Rhizobium than to the slow-growing species; (2) Although the
fast-growing PRC strains shared several microbiological and physiological
characteristics in common with other fast-growing species of Rhizobium,
they possessed some unique characteristics, such as the ability to utilize
ethanol as the sole source of carbon and energy and to hydrolyze gelatin; (3)
The taxonomic position of the fast-growing soybean rhizobia must logically be
in the new genus Rhizobium.
Based on their symbiotic characteristics, may warrant seperate species
status; (4) All of the fast-growing PRC soybean strains which were examined
could be separated into at least three somatic serological groups based on
immunofluorescence and immunodiffusion reactions. Immunodiffusion analysis of heat-labile whole-cell antigens
indicated that all the strains shared at least one heatlabile common antigen;
(5) All of the fast-growing PRC strains contained 1 - 4 high molecular weight
plasmids (M.W. > 100 Mdal). While
most of the strains shared plasmids with similar size, restriction endonuclease
profiles of plasmids from three of the strains were vastly different; (6) In
one of the strains, growth in the presence of subbacteriostatic levels of
acridine orange was effective in producing mutants cured of their largest
plasmid. In these mutants the loss of
the large plasmid led to the loss of nodulating ability indicating that
nodulation genes might be plasmid borne in this group of organisms. High-temperature curing of a smaller plasmid
in another strain did not lead to the loss of modulating ability or alteration
of symbiotic effectiveness on soybean cultivars; (7) Although one of the
fast-growing soybean strains was capable of receiving and maintaining the R.
leguminosarum pea host-range (Sym) plasmid, the resulting
transconjugants were unable to express the pea nodulation genes located on the
plasmid. On the otherhand, transfer of pJB5JI to
two R. trifolii strains resulted in pea-nodulatiog
transconjugants indicating that the expression of genes on the Sym plasmid
depends on the genetic backround it resides in; (8) Plasmid pJB5JI was differentially expressed in R. trifolii
transconjugants depending on whether transconjugants were made from Nod(#2)
or Nod+ (0403) R. trifolii strains. That is, R. trifolii #2
transconjugants were Nod+, Fix+ on peas, while R. trifolii
0403 transconjugants were Nod+, Fix- on peas; (9) In some
R. trifolii 0403 transconjugants, there was variable expression
of plasmid pJB5JI depending on whether
the transconjugants were isolated from culture or subsequently reisolated from
pea or clover nodules. Culture isolated
transconjugants were Nod+, Fix- on peas and Nod+, Fix+
on clover, while nodule reisolates were Nod+, Fix+; Nod+,
Fix-; or Nod- on peas; and (10) Transfer of plasmid pJB5JI
to R. trifolii 0403 resulted in
the construction of unique transconjugants which had the ability to nodulate
two legume hosts.
The fast-growing soybean
strains from China may provide an effective tool for a better understanding of
the genetics of the soybean-Rhizobium symbiosis since they appear more
amenable to genetic manipulations than the typical slow-growing soybean
symbiosis.
TABLE OF CONTENTS
ACKNOWLEDGEMENTS .................................. 3
ABSTRACT .......................................... 4
LIST OF TABLES .................................... 8
LIST OF FIGURES ................................... 10
PREFACE ........................................... 12
CHAPTER I. INTRODUCTION
AND OBJECTIVES ........... 13
CHAPTER II. LITERATURE
SURVEY..................... 15
CHAPTER
III. BIOCHEMICAL AND PHYSIOLOGICAL
RELATEDNESS
OF FAST-GROWING
SOYBEAN RHIZOBIA TO OTHER
FAST- AND SLOW-GROWING RHIZOBIA ..... 33
CHAPTER
IV. PLASMIDS OF FAST-GROWING SOYBEAN
RHIZOBIA: PROFILES, RESTRICTION
ENDONUCLEASE PATTERNS, AND ROLE
IN NODULATION ........................ 77
CHAPTER
V. SEROLOGICAL RELATEDNESS OF FAST-
GROWING SOYBEAN RHIZOBIA TO OTHER
FAST- AND SLOW-GROWING RHIZOBIA ....... 118
CHAPTER VI. GENERAL
SUMMARY ...................... 133
LITERATURE CITED .................................. 135
LIST
OF TABLES
Table Page
I-1 General
characteristics of fast- and
slow-growing rhizobia
.............................. 20
I-2 Current and
proposed classification of
of the root-nodule
bacteria ........................ 29
III-1 Mean generation
times and final pH
of the medium of several
fast- and slow-
growing rhizobia when
grown in various
media
..............................................
45
III-2 Mean generation
times of fast- and
slow-growing soybean rhizobia
in
sterile soil
.......................................
47
III-3 Biochemical
characteristics of fast-
and slow-growing soybean
rhizobia .................. 48
III-4 Litmus milk
reactions of fast- and
slow-growing soybean
rhizobia ...................... 50
III-5 Carbohydrate
utilization by fast- and
slow-growing soybean
rhizobia ...................... 52
III-6 Growth
responses of fast- and slow-
growing soybean rhizobia
to various
antibiotics
........................................
53
III-7 Enzyme
activities of fast- and slow-
growing soybean rhizobia
........................... 54
III-8 Growth of fast-
and slow-growing
rhizobia in ethanol
................................ 56
III-9 Growth and
survival of fast- and
slow-growing rhizobia in
ethanol ................... 57
III-10 Growth parameters of fast-growing
strain USDA 191 in several
concentrations of ethanol ......................... 59
LIST OF TABLES (CONTINUED)
IV-1 Bacteria used and their sources
................... 83
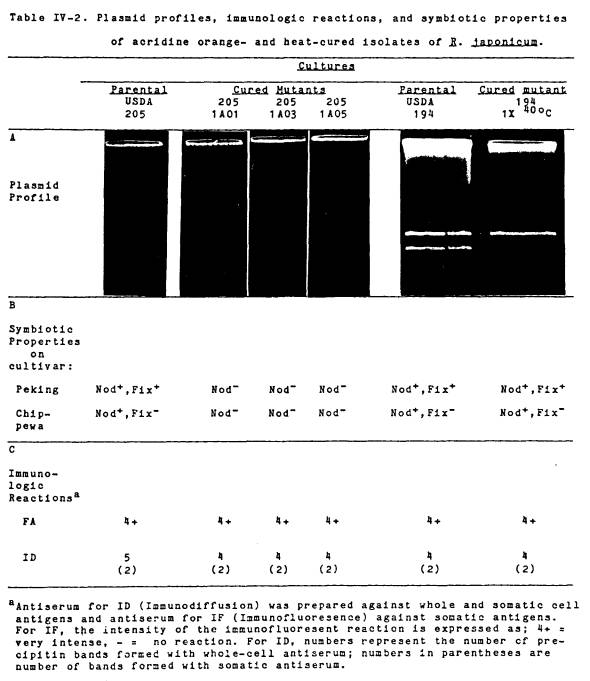
IV-2 Plasmid
profiles, immunologic
reactions,
and symbiotic
properties
of acridine orange-
and
heat-cured R. japonicum ....................... 96
IV-3 Frequency
of natural kanamycin
resistance
in several fast-
growing
R. japonicum and
R.
trifolii isolates .............................. 98
IV-4 Frequency
of transfer of plasmid
marker
in crosses between R.
leguminosarum,
R. japonicum,
R.
trifolii, and their
transconjugants
................................... 100
IV-5 Symbiotic
properties of R.
japonicum,
R. leguminosarum,
R.
trifolii, and their
Transconjugants
................................... 106
IV-6 Symbiotic
properties of R.
trifolii
transconjugants
reisolated
from nodules ...........................
111
V-1 Bacteria used and their sources
................... 122
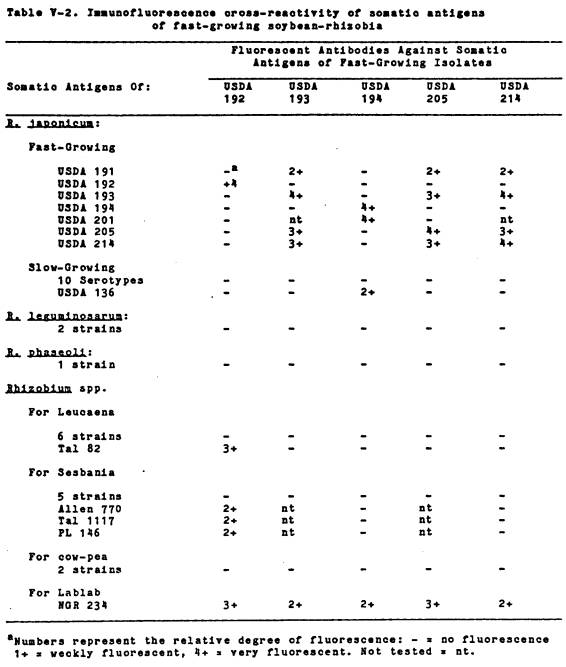
V-2 Immunofluorescence
cross-
reactivity
of somatic antigens
of
fast-growing soybean rhizobia .................. 126
V-3 Immunodiffusion
analysis of fast
and
slow-growing rhizobia ......................... 129
LIST OF FIGURES
Figure Page
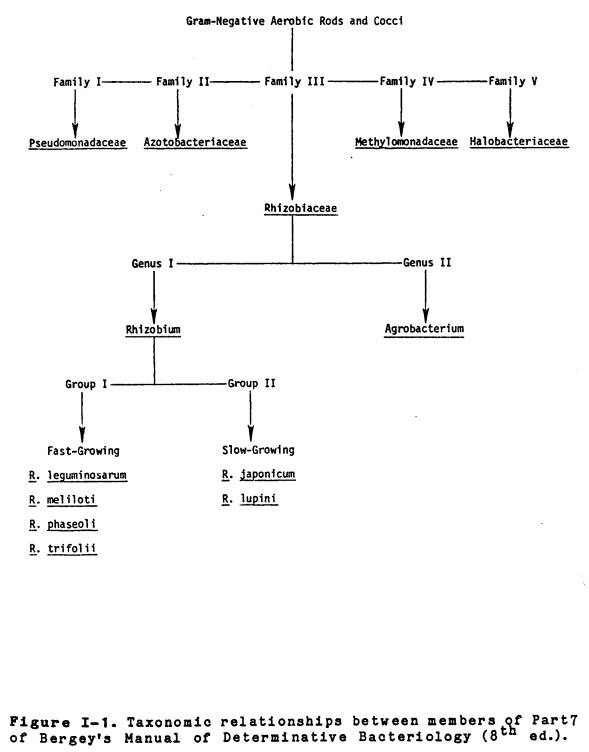
I-1 Taxonomic
relationships between
members
of part 7 of Bergey’s
Manual
of Determinative
Bacteriology
(8th edition) ........................
16
III-1 Growth
and ethanol utilization
of
USDA 191 in Bishop’s medium
with
various concentrations
of
ethanol ........................................ 61
III-2 Ethanol
concentration step-up
USDA
191 while growing on two
concentrations
of ethanol ......................... 63
III-3 Relationships
between inoculum
size
and lag phase of USDA 191
when
growing in 0.4% ethanol ......................
65
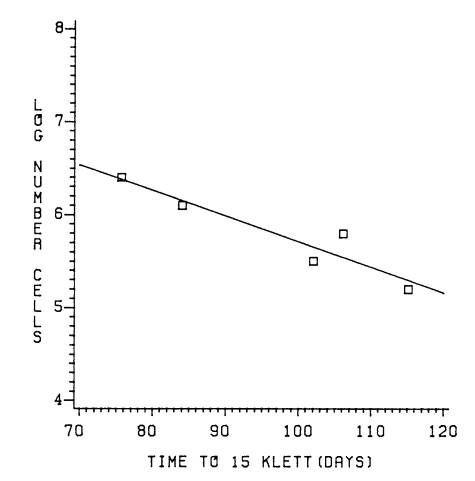
III-4 Regression
line of time to
reach
15 Klett units and log
number
of inoculum size ...........................
67
III-5 Growth and
mannitol and ethanol
utilization of USDA 191
........................... 69
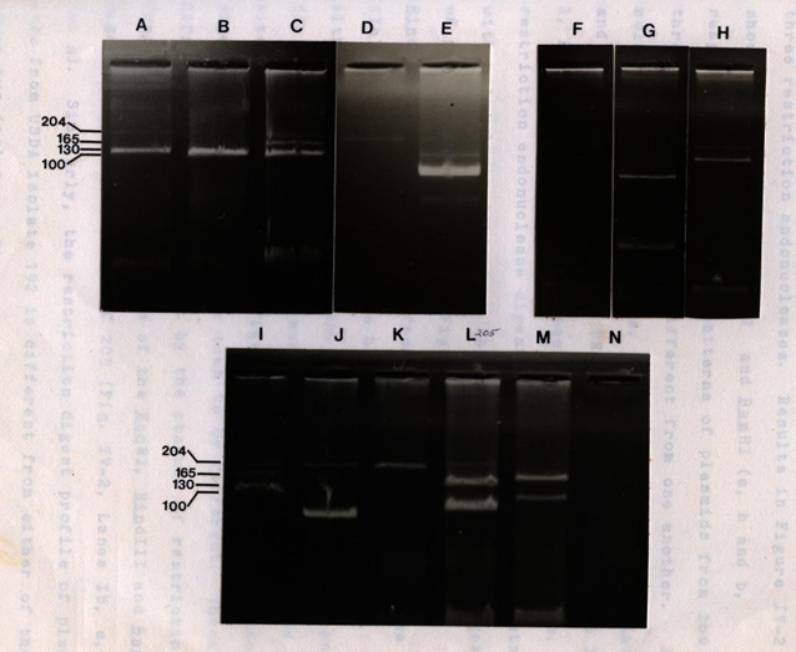
IV-1 Agarose gel
electrophoresis of
plasmid DNA from fast-
and
slow-growing rhizobia
............................. 91
IV-2 Restriction
endonuclease digest
of plasmids from the
fast-growing
PRC R.
japonicum .................................. 94
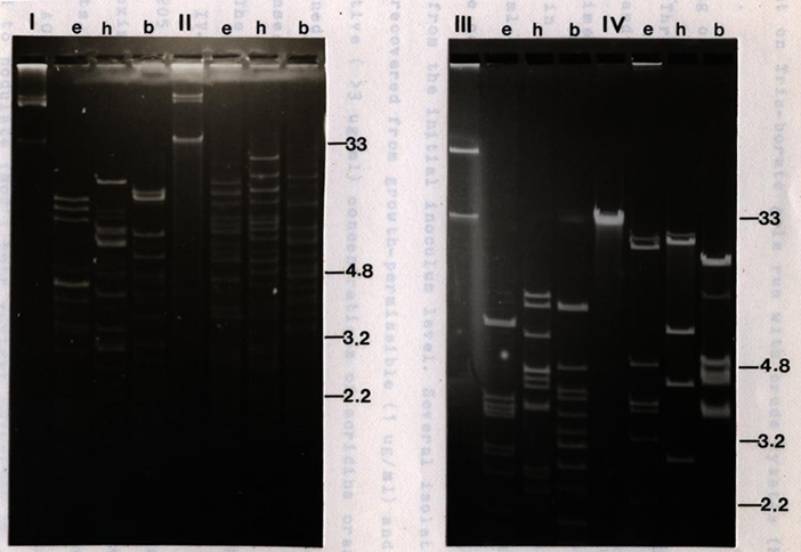
IV-3 Plasmid profiles
of R. japonicum
USDA
201 and its transconjugants ..................
103
IV-4 Plasmid profiles
of R. leguminosarum
6015
and its transconjugants ......................
105
IV-5 Plasmid
profiles of R. trifolii
strains
#2 and 0403 and their
transconjugants
................................... 109
LIST
OF FIGURES (CONTINUED)
V-1 Serological
analysis of several
fast-growing
soybean rhizobia .....................
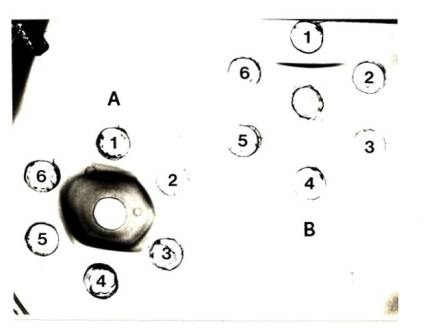
128
PREFACE
Some material presented
in this dissertation was obtained with the help and collaboration of several
investigators. I would specifically
like to acknowledge Dr. Harold Keyser for his collaboration on several biochemical
tests presented in Chapter III. I would
also like to acknowledge Stephen Dowdle’s contributions to growth rate data
presented in Chapter III. Lastly, I
wish to acknowledge Heidii Fugii for her contributions to R. trifolii
plasmid transfer experiments presented in Chapter IV and Ben Bohlool for the
preparation of antibodies and immunofluorescence results.
CHAPTER
I
INTRODUCTION
AND OBJECTIVES
Recent technological
advances in molecular genetics have prompted renewed interest in the genetics
of nitrogen fixation. Although many organisms
are capable of fixing atmospheric dinitrogen into plant-useable nitrogen
sources, a large number of investigators have focused their attention on
members of the genus Rhizobium.
This group of microorganisms is unique in that the fixation of nitrogen
only occurs when the rhizobia are in a symbiotic state with the plant host.
The "typical"
slow-growing rhizobia that form nodules on the roots of soybeans, Glycine
max, have now been reclassified in the new genus Bradyrhizobium (79). Recently, Keyser et al. (84)
reported the isolation of fast-growing soybean-rhizobia from nodules and soil
collected in the People’s Republic of China (PRC). All of the strains they examined (isolated in the Honan,
Shantung, and Sanshi Provinces of China) had mean generation times (in yeast
extract mannitol medium) between 2 and 5 hours and lowered the pH of the
culture medium (final pH’s ranged from 4.3-6.7). All formed effective symbioses with wild soybeans (Glycine
soja) and an unbred cultivar from China (Glycine Max, cultivar
Peking), but formed ineffective symbioses with most commercial soybean
cultivars. It should be noted that Glycine soja is reported
to be the putative wild ancestor of todays cultivated soybeans (55). In addition, the isolates were reported (84)
to ineffectively nodulate Macroptilium atropurpurum, Macroptilium
laythyroides and Sesbania cannabina, all of which are
promiscuous legumes which are nodulated by a wide variety of rhizobia. However, the PRC isolates did not nodulate, Leucaena
leucocephala (Hale Koa), Medicago sativa (alfalfa), Trifolium
repens (clover) or Astragalus sinicus.
The genetics of the root
nodule bacteria of such an important crop as soybeans has been neglected,
perhaps due to difficulties in consistently demonstrating plasmids in all the
slow-growing B. japonicum strains. Furthermore, the location of symbiosis-related genes in
slow-growing rhizobia is largely unknown.
Although several investigators (83,95,113) have transferred plasmids to
slow-growing rhizobia, in all instances, the plasmids used were the P1
incompatibility group plasmids (originally from Pseudomonas aeruginosa). Also, not all of the plasmids used could be
transferred to all of the strains investigated.
The objectives of this
dissertation were to determine: 1, the degree of relatedness of the
fast-growing soybean rhizobia to each other, to the “typical” slow-growing B.
japonicum, and to other fast-growing rhizobia. Relationships between the PRC rhizobia and
other fast- and slow-growing rhizobia were assessed at the biochemical and
physiological levels in order to ascertain the relative taxonomic position of
these newly described isolates; 2, the serological relationships of the
fast-growing soybean rhizobia to other fast- and slow-growing rhizobia; 3, whether
the fast-growing soybean rhizobia contain large molecular weight plasmids and
if plasmids were present, whether there are structural relationships between
plasmids from different strains; 4, whether symbiosis-related genes are
plasmid-borne in these isolates; and 5, whether the fast-growing soybean
rhizobia were capable of accepting, maintaining, and expressing plasmids from
other fast-growing rhizobia.
CHAPTER
II
LITERATURE SURVEY
Members of the genus Rhizobium can best be described as a
heterogeneous group of Gram-negative, aerobic, heterotrophic, non-sporeforming
rods which have the ability to invade and form nodules on the roots of
leguminous plants (80).
Within these root-nodules, a symbiotic state exists between the
rhizobial partner (microsymbiont) and the plant partner (macrosymbiont). One result of this symbiosis is the fixation
of atmospheric dinitrogen into ammonia by the rhizobial partner, in exchange
for protection and a source of photosynthetically fixed carbon, provided by the
host plant. Although the process of
nitrogen fixation (via the enzymecomplex nitrogenase) is not solely restricted
to members of the genus Rhizobium, it is the root-nodule-symbiosis with
plants of the family Leguminosae, which is almost exclusively a
property of this group of microorganisms (133).
The family Rhizobiaceae
is currently listed (80) along with Pseudomonadaceae, Azotobacteriaceae
Methylmonadaceae, and Halobacteriaceae in part 7 of the 8th
edition of Bergey’s Manual of Determinative Bacteriology (See Figure I-1 for
relationships between families and genera).
The only major feature shared by all these families is their aerobic
metabolism and negative Gram-reaction, however, members of the Azotobacteriaceae
have the ability to fix atmospheric dinitrogen in the free-living state. The Rhizobiaceae are divided into two
genera, Rhizobium and Agrobacterium, with the major
distinguishing characteristic between the two groups being the ability of Agrobacterium
(with the exception of A. radiobacter) to form cortical
hypertrophies (Crown Galls) on the roots and stems of numerous dicotyledonous
plants. The current classification
scheme bases species
|
|
designation within the
genus Rhizobium exclusively on the ability of a specific bacterium to
nodulate a given legume host. This
approach divides rhizobia into several “plant-infection” groups and results in
the designation of six species: Rhizobium japonicum (Soybean), R.
leguminosarum (Pea), R. lupini (Lupen), R. meliloti
(Alfalfa), R. phaseoli (Bean), and R.
trifolii (Clover).
Although the division of
rhizobia into “plant-infection” groups does have certain practical
applications, there are also some major problems with this scheme
(133,138). While some rhizobial species
groups may be correctly defined by the relatively small numbers of legumes
they are able to nodulate, others can nodulate a great number of different
legumes. For example, R. meliloti
isolates which can nodulate most species of Medicago (Alfalfa) and Melilotus
(Sweet Clover) but not Phaseolus (Beans), Trifolium (Clover), or Viccia
(Vetch), are said to have cross-inoculation group specificity. On the other hand, some isolates of R.
leguminosarum, in addition to forming nodules on Pisum (Peas),
are able to nodulate Viecia, Lathyrus (Roughpea, Grasspea), and Lens
(Lentils) (138). It should be noted
however, that the ability to form nodules on the roots of leguminous plants is a
property contributed to by both the micro- and macrosymbiont. In addition to the above mentioned
shortcomings, there exists additional problems in using schemes based on
plant-infection groupings. The first
problem is that there are many rhizobia that do not fit into the present scheme
(they have other plant host requirements) and are thus placed into separate “catch-all”
groups. One such example is the
so-called “cowpea-miscellany” group.
Members of this group have the ability to nodulate a wide variety of
host plants, such as, Vigna (Cowpea), Glycine (Soybeans), and Macroptilium (Siratro). Another example of a “catch-all” group are
the lotus rhizobia (73) which have the ability to form nodules on legumes of
the genera Lupinus (Lupine), Ornithopus (Serradella), Anthyllis
(Kidney vetch), and Astragalus (Tragacanth). Another problem with the plant-infection scheme involves those
organisms which are no longer capable of effectively modulating their
designated host plant. For example, by definition, those organisms able to
nodulate clover are referred to as R. trifolii, however, R.
trifolii mutants (both spontaneous and induced) are available
which have been rendered non-nodulating.
By the above definition, they should no longer be referred to as
rhizobia.
A. Fast- and slow-growing rhizobia
The genus Rhizobium
can also be divided into two major groups depending upon growth rates and
effects of growth on the pH of yeast extract mannitol (YEM) culture medium (1,41,53,73.79,80,97,106,133,138).
The
"fast-growing" rhizobia have mean generation times between two and
four hours and produce a net decrease in the pH of YEM culture medium. On the other hand, those rhizobia referred
to as "slow-growing" have mean generation times of six hours and
longer and do not lower the pH of the medium (133).
Generally
speaking, R. leguminosarum, R. meliloti, R. phaseoli,
R. trifolii, and Rhizobium spp. capable of nodulating Leucaena
and Sesbania are characterized as fast-growing and acid-producing,
while R. japonicum, R. lupini,
and the "cowpea group" are characterized as slow-growing and
alkaline-producing. As was pointed out
by Vincent (133),
the lotus
rhizobia present some taxonomic difficulties, since there are both fast- and
slow-growing strains which have similar host requirements. Similarly, among the
Rhizobium lupini, both fast- and slow-growing strains can be
isolated. More recently, Keyser et
al. (84) reported
that in addition to the slow-growers, there are also fast-growing rhizobia able
to nodulate soybeans.
B. Biochemical attributes of rhizobia
Consistent with the
division of rhizobia into fast- and slow-growing groups based on their growth
rate and effect on the pH of YEM culture medium, is their division into two
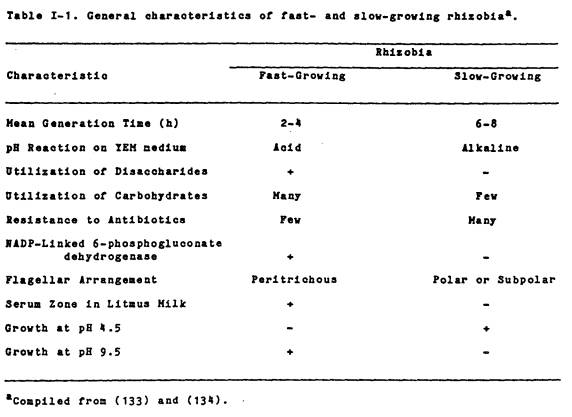
broad groups on the basis of several other characteristics (Table I-1). Graham and Parker (53) have indicated that
the fast-growing rhizobia may be differentiated from the slow-growing ones by
the presence of methylene blue-staining cytoplasmic granules and by colony size
on YEM agar. The former have such
granules and produce much larger colonies than the latter. As was pointed out by Fred et al.
(44), Graham and Parker (53), and Vincent (133), fast-growing rhizobia tend to
utilize a wider variety of carbohydrates than do the slow-growers. Although the utilization of a specific sugar
is in itself not a useful tool for differentiating rhizobia, a clear difference
does exist in the pattern of utilization of a large number of carbohydrates by
fast- and slow-growing strains. The
fast-growers also generally have the ability to utilize disaccharides as a sole
source of carbon and energy for growth.
Glenn and Dilworth (48), Graham (50), and Martinez-De Drets and Arias
(101) pointed out that slow-growing rhizobia are generally unable to utilize
disaccharides. Glenn and Dilworth (48)
indicated that the slow-growers apparently lack both uptake systems and catabolic
enzymes for disaccharide utilization.
For example, fast-growing rhizobia have B-galactosidase activity, while
the slow-growers lack this enzyme.
Martinez- De Drets and Arias (101,102) have also shown that although
both fast- and slow-growing rhizobia have NAD-linked 6-phosphogluconate
activity (6-PGA), only the fast-growers have NADP-linked 6PGA. Thus, rhizobia can also be separated into
fast- and slow-growing groups based on the presence or absence of enzymes of
the pentose phosphate pathway.
Fast- and slow-growing
rhizobia can also be separated on the basis
|
|
of their relative
tolerance to pH and NaCl. Previous
studies of rhizobia (20,43,53,106) showed that fast-growers were relatively
more alkali-tolerant and acid-sensitive, than the slow-growers. Also, Graham and Parker (53) showed that
among rhizobia, tolerance to 2% NaCl was restricted to the fast-growing R.
meliloti. The responses of
rhizobia in litmus milk (44,53) and their relative resistance to antibiotics
(128) have also been used to separate rhizobia into two growth-rate groups.
Fast-growers tend to produce acid and peptonization reactions in litmus milk
and are generally sensitive to antibiotics.
The slow-growers on the other hand, do not possess these
characteristics.
C. Deoxyribonucleic acid base ratios and
homology
Rhizobia can also be
separated into several groups based on DNA base ratios and nucleic acid
hybridizations. While DNA base ratios,
mole percent G+C, for Rhizobium are relatively broad ranged [values
ranging from 59-66% (32)], they do indicate relationships between strains. De Ley and Rassel (32) indicated that the
peritrichously flagellated fast-growing rhizobia, R. leguminosarum,
R. phaseoli, and R. trifolii,
tend to have low %G+C values, ranging from 59-63%, while the polar to subpolarly flagellated
slow-growers, R. japonicum, R. lupini, and some of the cowpea
miscellany, have relatively high %G+C values, 63-65%.
Heberlein
et al. (57) reported
that with the exception of Agrobacterium pseudotsugae and R.
japonicum, the agrobacteria and rhizobia which they examined had about
the same %G+C content, about 59-63%. Rhizobium japonicum on the
other hand, had G+C values of about 64-65%. However, Elkan (40) examined 25 strains of R. japonicum
and found their mole percentage G+C to be relatively homogeneous, 61-64%. Elkan (40) also indicated that although %G+C values are
significantly different between some strains of R. japonicum, these values fall within the range of other
fast- and slow-growing rhizobia and thus, are of limited taxonomic value.
DNA-DNA homologies have
been used by several investigators (57,47,64) to study the degree of genetic relatedness between strains
and isolates of Rhizobium. The
homology studies of Gibbons and Gregory (47) and Heberlein et al. (57) have indicated that there
is a sharp line of demarcation between the fast-growing species (R. leguminosarum,
R. trifolii, R. phaseoli, and R. meliloti) and the
slow-growing ones (R. japonicum and R. lupini). Hollis et al. (64) have
recently shown that the slow-growing strains of R. japonicum which they examined could be separated into at least three distinct DNA
homology groups. These authors also
indicated that there was little homology between DNA from several R. japonicum strains and R. leguminosarum, R. meliloti, R.
phaseoli, R. trifolii, and Agrobacterium tumefaciens. However, there was substantial homology between the slow-growing
R. lupini and R. japonicum. Similarly, Crowe et al.
(29) showed that the 113 fast-growing
and 9 slow-growing strains of rhizobia which they examined could be placed into
4 DNA homology groups, and that the fast-growers were clearly separated from
the slow-growers. Interestingly, these
authors also indicated that there was little homology between fast- and slow-growing
lotus rhizobia. However, the two groups
were more closely related to each other than to the other fast- and
slow-growing rhizobia which were examined.
Thus, fast- and slow-growing rhizobia can also be separated into two
groups on the basis of DNA-DNA homologies.
In addition to the above
mentioned characteristics, fast- and slow-growing rhizobia have also been
separated into fast- and slow-growing groups on the basis of polyacrylamide
gel electrophoresis of cell proteins (119) and nodule-bacteroid inclusion
bodies (28).
D. Serological relationships
Serological techniques
have been used routinely in the study of rhizobia: 1, to obtain information
about their antigenic composition (52,68); 2, for strain identification (13,78,104,123); 3, to investigate the
serological relatedness of strains and species of Rhizobium (36,37,46,69,71,136,137) and 4, for ecological studies (13,14,15,16,104,85,121,122,123,). Three techniques in particular have found wide acceptance
in serological investigations of the Rhizobiaceae: agglutination,
immunodiffusion (Ouchterlony Gel Diffusion), and immunofluorescence.
Agglutination techniques
were the first of the serological methods applied to the study of rhizobia
(38). In general, early serological
studies were concerned mainly with the relationships between serological
groupings and host-specificity. Results
of most studies indicated that rhizobia are a serologically heterogeneous group
of organisms. Stevens (127) and Wright (140) found that different
strains isolated from the same host-plant species could be serologically
unrelated. Hughes and Vincent (67) found that even strains
isolated from different nodules on the same plant could be unrelated
serologically. However, in their study,
Bushnell and Sarles (23) found
that some strains from different cross-inoculation groups were serologically
related. Kleczkowski and Thornton (86)
indicated that the ability of a rhizobial strain to nodulate a particular
host-plant is not necessarily related to it serological characteristics as
detected by agglutinations. Bushnell
and Sarles (23) examined rhizobia from the soybean, cowpea, and lupin
cross-inoculation groups and found no correlation between the ability of
certain strains in one group to cross-inoculate another, and their ability to
cross-agglutinate. In addition, Stevens
(127) and Bushnell and Sarles (23) indicated that due to their serologic heterogeneity,
all strains within the same Rhizobium species cannot be identified by
agglutination reactions with a limited number of antisera.
Despite the inability to
show correlations between host-specificity and serological groupings,
agglutination reactions have been used to assess the serological relatedness
of strains and species of Rhizobium. Kleczkowski and Thornton
(86), using whole-cell antisera against 4 strains of R. trifolii
and 2 strains of R. leguminosarum, examined the agglutination
cross-reactions of antigens from 161 strains of R. trifolii, 29 R.
leguminosarum, 5 strains of R. meliloti and R. lupini,
and 13 non-Rhizobium soil isolates.
Results of their study indicated that while no cross-reactions occurred
outside of the clover and pea groups and with the 13 soil isolates, some
cross-reactions occurred between the two groups. And, while some antisera were quite specific, others were
relatively non-specific.
In their studies on the
serological relationships of 25 strains of the slow-growing R. japonicum,
Koontz and Faber (88) identified 6 somatic serogroups using agglutination adsorption
reactions. Wright et al.
(140) similarly found 6 serogroups among the R. japonicum
they examined, however, these authors did not differentiate between somatic and
flagellar antigens. Date and Decker
(30) analyzed 28 strains of R. japonicum and found 17 somatic
serogroups on the basis of cross-reactions and agglutination cross-adsorptions.
Graham (49) tested 113
strains of Rhizobium for agglutination by whole cell antisera produced
against 58 strains of Rhizobium and 16 Agrobacterium strains. The
results of his study of whole- and somatic-cell antigens indicated that the
rhizobia could be separated into three serologically distinct groups: 1, R.
trifolii, R. leguminosarum, and R. phaseoli;
2, R. japonicum, R. lupini, and Rhizobium
spp. of the cowpea miscellany; and 3, R. meliloti. While there were no cross-reactions between
the groups, there were cross-reactions within the groups. Graham (49) also indicated that strains of R.
meliloti showed some serological relatedness to Agrobacterium tumefaciens
and A. radiobacter and that agglutination cross-reactions were
greater with whole-cell antigens than with somatic-cell antigens.
Immunodiffusion
techniques, specifically Ouchterlony double-diffusion, have also been used
extensively to investigate the serological relationships between strains and
species of Rhizobium (36,37,51,69,70,71,125). The technique relies on the separation of soluble, diffusible
antigens through an agar-gel matrix.
Relationships between various antigens and antisera are determined by examining the nature
of the interaction at the junction of precipitin bands from various wells. Gel diffusion methods have been used in the
study of rhizobia because they permit the rapid enumeration of soluble
antigens, the techniques are relatively simple, and they can be used to study
serological relationships of strains at the single antigen level. Dudman (37) has indicated that while
agglutination reactions can be used to separate rhizobia into serological
groups, agglutination techniques lack the resolving power of immunodiffusion in
distinguishing between antigenically identical and closely related, but not
identical strains.
Dudman (36) was the first
investigator to use immunodiffusion techniques to study the serological
relatedness of strains and species of Rhizobium. In his study of the extracellular soluble
antigens of 2 strains of R. meliloti, Dudman (36) found that the
two strains examined shared all antigens accept several fast-moving ones. He proposed that since the strains did not
cross-agglutinate, that these strain-specific antigens could be used for
identification purposes.
Using gel
immunodiffusion, Skrdleta (125) divided the 11 slow-growing R. japonicum
which he examined into two basic somatic serogroups. While he detected the same
serogroups using agglutination reactions, he found that immunodiffusion
allowed him to show serological relationships between strains that were not
agglutinated by the same antisera.
Skrdleta (125) also
indicated that the somatic antigens were more specific than flagellar ones in
differentiating individual strains.
Dudman (37) in his study of seven
strains of R. japonicum found that pretreatment of antigens (by
boiling or ultrasonic disruption) was required for the proper immunodiffusion
analysis of these slow-growing strains.
Gibbins (46) found that while ultrasonic disruption prevented the
formation of precipitin bands in immunodiffusion reactions, band formation
could be restored by heating the sonicated antigen preparations.
While the use of somatic
antigens (heat-stable antigens) have been more specific than flagellar ones
(heat-labile) in differentiating individual strains of Rhizobium,
internal antigens have also been reported (135) to provide some insight into the serological relatedness
of fast- and slow-growing rhizobia.
Using whole-cell antisera against three strains of R. japonicum,
Vincent et al. (137) studied the internal
antigens of sixty-nine strains of Rhizobium and 5 Agrobacterium strains.
Immunodiffusion reactions
indicated that at least one common antigen was present in 13 strains of R. japonicum,
4 strains of R. lupini, and 4 strains of the slow-growing cowpea and
lotus rhizobia. Their results also indicated that the forty-six fast-growing
rhizobia examined were readily distinguished from the slow-growing strains and
that the 5 strains of agrobacteria grouped with the fast-growing rhizobia. More recently, Pankhurst (110) studied the
immunodiffusion cross-reactions of somatic and interal antigens from 62 fast-
and slow-growing strains of lotus rhizobia. Results of his study indicated that
while the fast- and slow-growers shared no common somatic antigens, internal
antigens were shared by all of the fastgrowing strains, and with seven
exceptions, by all of the slow-growing strains.
The fluorescent antibody
technique is the method of choice for the direct examination and identification
of strains of rhizobia in culture and nodules (13,14,123) and for the
enumeration of specific strains directly in soi1 (14,85,122). The major advantages of immunofluorescence
over other techniques is that only small amounts of antigen and antibody are
needed (121), the procedures are relatively rapid, and its the only technique
readily applicable to the study of rhizobia in situ (16).
Vincent (135) in his
recent review of the literature has pointed out that when serological (137) and
other taxonomic evidence [see (73) and (79)] are taken together, clear
relationships among rhizobia can be recognized. That is; 1, there is a closer relationship between the
fast-growing species of Rhizobium and Agrobacterium than there is
between the fast- and slow-growing groups of rhizobia; 2, R. trifolii
and R. phaseoli should be made separate biovars of the species R.
leguminosarum; 3, R. meliloti is so different from
other species of Rhizobium, that it requires its own species status; 4,
among the slow-growers, the slow-growing soybean rhizobia should remain as a
separate species group and be included in the new genus Bradyrhizobium;
and 5, the fast- and slow-growing lotus rhizobia fall within the genera Rhizobium
and Bradyrhizobium, respectively.
E. Current Rhizobium taxonomy
Due to the large number
of differences existing between the fast- and slow-growing rhizobia, and to
inadequacies in the cross-inoculation-plant-infection group scheme, a new
classification scheme for rhizobia has been proposed [see Jordan (79) and
Jarvis (73)]. This scheme (since
adopted by the International Subcommittee on Agrobacterium and Rhizobium)
divides rhizobia taxonomically into fast- and slow-growing groups. The first group consists of the fast-growing
rhizobia. Those organisms previously
designated as R. leguminosarum, R. trifolii, and R.
phaseoli will be combined as one species, R.
leguminosarum (Table I-2), comprising three biovars (trifolii,
phaseoli, and viceae). Rhizobium
meliloti [which differs significantly from other rhizobia (134)], is
kept as a separate species group. The
slow-growing rhizobia, R. japonicum, were transferred to a separate genus, Bradyrhizobium. Only one species is present in this genus, R.
japonicum. Other slow-growing
rhizobia are to be referred to as Bradyrhizobium spp., with the name of
the designated-nodulated plant following in parentheses. In the remainder of this dissertation, those
slow-growing organisms previously referred to as Rhizobium japonicum
will be referred to as Bradyrhizobium japonicum and R.
japonicum will be used only to refer to the fast-growing
soybean-rhizobia from China. It should
be noted that the slow-growing rhizobia formerly referred to as R. lupini
are not included as a separate species in the new genus Bradyrhizobium,
since their only major distinguishing characteristic is their nodulation
affinity for Ornithopus and Lupinus. Subsequent to the
adoption of this new scheme (which will be appearing in the forthcoming edition
of Bergey’s Manual of Determinative Bacteriology), Jarvis et al.
(73) have proposed that another
species group be included in the new genus Rhizobium. This species, Rhizobium loti refers
to the heterogeneous group of fast-growing rhizobia which effectively nodulate
Lotus corniculatus (birds-foot treefoil), Lupinus densiflorus
(lupines) and Anthyllis vulneraria (kidney vetch). It should be noted however, that some
strains of the lotus rhizobia ineffectively nodulate a great variety of
platns. In addition, the slow-growing
lotus rhizobia will be included in the new genus Bradyrhizobium.
|
|
F. Genetics of the Rhizobium-legume
symbiosis
The study of the genetics
of the root-nodule bacteria had its beginning in the 1960’s (91). It has been the general interest of many
investigators to study those genes necessary for nodulation and nitrogen
fixation. Techniques used to study the genetics of the Rhizobium symbiosis
have included mutagenesis, transduction, transformation, and conjugation. More recently, the genetic aspects of the Rhizobium-legume
symbiosis have been investigated using molecular genetic techniques.
Although Beringer (8) has
indicated that transformation studies with Rhizobium may have begun in
the 1940’s by Krasilnikov (92,93),
he has also pointed out that this work cannot adequately be evaluated.
Initially, transduction, transformation, and conjugation were explored in order
to facilitate gene transfer between rhizobia.
Although transduction has been used by Kowalski (91) and Buchanon-Wollaston
(21) to transfer DNA between strains of R. meliloti and between R.
leguminosarum and R. trifolii, its general application in
the study of rhizobia has been limited.
This may be due to the fact that only small segments of DNA can be
transferred and that the available phages only mediated generalized
transduction (8). While transformation
systems have been developed for several fast and slow-growing rhizobia
(2,115,116,117), the selected markers in most cases have been streptomycin
resistance, gelatinase activity, or the conversion of amino acid-auxotrophs to
prototrophs. However, in 1978, Page (109) transformed a strain of
the free-living nitrogen fixing organism, Azotobacter vinelandii, which
was unable to fix nitrogen (Nif-), to a Nif+ phenotype,
using DNA isolated from several rhizobia.
Conjugation within and
between Rhizobium species has been the most promising system for gene
transfer. Initial studies centered
around the construction of chromosomal linkage maps (4,6,7,59,87). In most instances, the P1 group plasmids
from Pseudomonas aeruginosa (RP4, R68, and R68.45) were used to
mobilize chromosomal DNA.
Mutants defective in
symbiosis-related function are essential for most genetic studies. Three approaches were taken to construct
nodulation (Nod) and nitrogen fixation (Nif) deficient strains. The first approach, used by Maier and Brill
(99) and Beringer (5), utilized "conventional" chemical mutagenesis
(usually N-methyl-N’-nitro-N-nitrosoguanidine). The second was that used by Beringer et al. (6) in
which the mutagenic agents were transposable genetic elements (transposons).
Transposons (TN) can integrate into many sites within the chromosomes of R. leguminosarum, R. trifolii,
and R. phaseoli (6) and into the indigenous plasmids of R. leguminosarum (76) and in doing so,
result in the production of Nif- and Nod- mutants. In the last approach, used by Zurkowski and
Lorkiewicz (142), Casse, (25), and Higashi (60), nodulation deficient R. trifolii and R. leguminosarum mutants were obtained
following acridine orange and heat curing of indigenous plasmids.
G. Rhizobium Plasmids
As was indicated above,
mutagenic agents, in addition to causing chromosomal mutations, can also cause
mutations in plasmid DNA. At the
present time, most of the fast-growing rhizobia that have been examined have
been shown to contain large [ M.W. > 100 megadaltons (Mdal)] plasmids
(17,18,25,54,66,94,107,114,120).
Initially, these large plasmids were only infrequently detected in
rhizobia. This was most likely due to
the use of techniques (such as the cleared-lysate method) designed for the
isolation of low M.W. plasmids. In the
past few years, several plasmid-borne functions have been identified in some of
the fast-growing Rhizobium species.
These include; medium bacteriocin production (61), melanin production
(10), hydrogen uptake (HUP) (19), host-range specificity
(10,22,60,65,66,76,142) and nitrogenase components (39,108,120).
As was indicated by
Beringer (8), future genetic studies of Rhizobium will be dominated by
studies of plasmids. One group of
plasmids which have captured considerable interest among investigators are
referred to as the symbiotic (SYM) plasmids.
One such example is the R. leguminosarum SYM plasmid,
pRL1JI. This plasmid, modified by the
addition of the transposon TN5, (pRL1JI::TN5) (76) has been used by several
investigators (10,18,35,62,77) to transfer pea nodulation ability into R.
leguminosarum, R. phaseoli, and R. trifolii. This 130 Mdal plasmid carries some of the
nitrogen fixation (Nif) genes as well as genes for the nodulation of peas. Plasmid pRL1JI, originally shown by Hirsch
(61) to contain genes for medium bacteriocin production is a
self-transmissible (due to a segment of DNA referred to as Tra), conjugal
plasmid. However, the addition of TN5
to the plasmid has resulted in the loss of its expression of medium
bacteriocins (presumably due to transposition into the bacteriocin genes). The
addition of TN5 into the plasmid (now referred to as pJB5JI) has facilitated
its use in further studies, since the kanamycin resistance phenotype (donated
by TN5) of pJB5JI can be used to select for transconjugants on agar plates
containing the antibiotic. Due to its
transferability and selectability, the Rhizobium leguminosarum
symbiotic (SYM) plasmid, pJB5JI, allows for the examination of the functioning
of symbiosisrelated genes in various genetic backgrounds.
CHAPTER
III
BIOCHEMICAL
AND PHYSIOLOGICAL RELATEDNESS OF FAST-GROWING
SOYBEAN
RHIZOBIA TO OTHER FAST- AND SLOW-GROWING RHIZOBIA
Abstract
Fast-growing,
acid-producing soybean-rhizobia were examined to determine their degree of
biochemical and physiological relatedness to each other, to
"typical" slow-growing B. japonicum, and to other fast-growing
species of Rhizobium. While
both the fast- and slow-growing rhizobia were positive for catalase, urease,
oxidase, nitrate reductase, and penicillinase, the fast-growing R.
japonicum grouped with other fast-growing species of Rhizobium
in that they were tolerant to 2% NaCl, sensitive to a larger number of
antibiotics, capable of growth at pH 9.5, utilized a large variety of
carbohydrates (notably disaccharides), and produced serum zones in litmus
milk. In addition, these fastgrowing
strains were similar to other fast-growing species of Rhizobium in that
they had appreciable levels of B-galactosidase, NADP-linked 6-phosphogluconate
dehydrogenase, and had mean generation times much less than the typical
slow-growers when growing in culture media and in sterile soil. While the fast-growing soybean-rhizobia were
capable of substantial growth in ethanol, the typical fast-growers failed to
grow in ethanol and the slow-growers grew only poorly. Although the
fast-growing soybean-rhizobia share symbiotic host-specificity with the typical
slowgrowers, they appear biochemically and physiologically more closely
related to the other fast-growing species than to the "typical"
slow-growing B. japonicum.
Introduction
Species within the genus Rhizobium
have been divided into two groups (80,133) depending on their growth rate and effect on the pH of
yeast extract-mannitol (YEM) medium under standard laboratory conditions. The fast-growing rhizobia have mean
generation times of between two and four hours and produce a net decrease in
the pH of YEM culture medium, while those referred to as slow-growing have mean
generation times of six hours and longer and do not lower the pH of the medium
(133).
The typical slow-growers
that form nodules on the roots of soybeans (Glycine max) have in
the past been referred to as Rhizobium japonicum (80). Recently, these rhizobia have been
re-classified in a new genus, Bradyrhizobium, on the basis of their slow
growth rate and other characteristics (79) to distinguish them from fast-growing, acid-producing
root-nodule bacteria which now comprise the genus Rhizobium [see (73)].
Recently, Keyser et
al
(84) reported
the isolation of fast-growing soybean-rhizobia from root nodules and soil
collected in the provinces of Shansi, Honan, Shandong, and Shanghai in the
People’s Republic of China (PRC). The
isolates have been reported (84) to have mean doubling times between 2 and 4 hours and
lower the pH of YEM culture medium (final pH ranging from 4.7 to 6.7). All strains were reported to form effective
nitrogen-fixing nodules on wild perennial soybeans (Glycine soja)
and on an unbred soybean cultivar from China (cv Peking), but formed
ineffective symbioses with most commercial cultivars (84).
This report describes the
taxonomic investigation of the fast-growing, acid-producing PRC isolates. Several diagnostic microbiological,
physiological, and biochemical tests were performed in order to determine the
degree of relatedness of the fast-growing soybean rhizobia to each other, to
the "typical" slow-growing B. Japonicum, and to other
fast-growing rhizobia. It was assumed,
that in order to make any meaningful assessment of this newly described group
of organisms, that their relative taxonomic relationship to other rhizobia
needed to be determined.
Materials
and Methods
A. Bacterial Strains and Growth Conditions
The fast-growing
soybean-rhizobia, USDA 191, 192, 193, 194, 201, 205, 206, 208, 214, 217, and
257 were isolated from soil or nodules collected in the People’s Republic of
China [Keyser, et al. (84)].
The slow-growing Bradyrhizobium Japonicum, Chinese strains
PRC-005, 74, 113-2, 121-6, 2031, and B15 were obtained from T. S. Hu, Institute
of Soils and Fertilizers, Chinese Academy of Agricultural Sciences, Beijing,
People’s Republic of China. Bradyrhizobium
japonicum strains USDA 6, 31, 34, 74, 94, 110, 122, 123, 136, 138, 142,
and Y1, Yla, Y2, Y2a, Y3, K2, K2a, S1, S1a, were
from the USDA Culture Collection, Beltsville, Maryland. Rhizobium lupini
CC814s, NZP 2021, NZP 2238, SU 343, and NZP 2037, R.
leguminosarum Nitragin 92A3, R. phaseoli NZP 5097, NZP
5253, and NZP 5260, R. trifolii NZP 560, and WU 95, and R.
meliloti NZP 4013 were obtained from R. M. Greenwood, Department of
Scientific and Industrial Research, Palmerston North, New Zealand. Rhizobium leguminosarum HI 5-0
was isolated in Hawaii [May and Bohlool (104)]. Rhizobium leguminosarum 6015(pJB5JI) was obtained
from P. Hirsch, Max Planck Institute, Koln, FRG; Rhizobium phaseoli
Bel 7.1 from E. L. Schmidt, University of Minnesota, Minneapolis; Rhizobium
sp. (Leucaena) Tal-82, from the Niftal Project, Paia, Hawaii; and Rhizobium
sp. (Leucaena) UMKL 19 and R. leguminosarum
PRE from W. J. Broughton, Max
Planck Institute, Koln, FRG. Agrobacterium
tumefaciens 79 and 101 and Rhizobium sp. (Sesbania) 3F4a4
were from USDA Beltsville, Maryland. Rhizobium
meliloti L530 was obtained from B. Rolfe, Australia National University,
Canberra City, Australia. All Rhizobium
cultures were maintained on yeast extract-mannitol (YEM) agar slants of
the following composition in g/1: yeast extract, 1.0; mannitol, 10.0; K2HPO4·3H20,
0.65; MgSO4·7H20, 0.2; NaCl, 0.1; pH 6.9 (132). Agar slants used for the maintenance of
fast-growing rhizobia contained 0.05% CaCO3. Agrobacterium
cultures were maintained on Nutrient agar (Difco) slants. All cultures were incubated at 28-300C, subcultured at least once every
month and stored at 40C.
B. Staining, Morphology, and Cultural
Characteristics
Cultures were examined
for cell morphology and Gram reaction after 3 d of growth in YEM liquid
medium. Colony morphology was examined
on cultures grown for 6 d on YEM agar containing brom-thymol-blue, BTB,(0.25
mg/1). Motility was estimated from both
YEM liquid and agar cultures and on B5 agar medium (45). Cell size determinations were performed
using a calibrated ocular micrometer.
Fast- and slow-growing PRC isolates were identified using strain
specific fluorescent antibodies prepared according to Schmidt et al.
(123).
C. Biochemical Tests
Tolerance to pH extremes
was determined by inoculating 107 cells/ml from exponentially
growing YEM liquid cultures into tubes containing 10 ml of YEM liquid medium
which were adjusted to pH 4.5, 9.0, and 9.5.
Tubes were incubated at 300C for 14 d and scored for
growth. Tests were performed in
triplicate.
Tolerance to sodium
chloride was determined on YEM agar plates containing 2.0% NaCl. Plates were spread with 108
cells, and growth was scored after 14 d of incubation at 300C. Tests were done in triplicate.
For growth reactions in
litmus milk (Difco), tubes (10 ml/tube) were incubated in quadruplicate for 6
weeks at 300C and were examined for pH changes, reduction of litmus, and
peptonization (serum zone formation).
Production of
3-ketolactose was determined according to Bernaerts and De Ley (9). Agrobacterium tumefaciens was
used as a positive control for this test.
For gelatinase activity,
exponential phase cultures from YEM liquid medium were swabbed onto the surface
of tryptone yeast extract (TY) agar plates (62) containing 0.4% (w/v) gelatin
(Difco). Plates were incubated at 280C
for 7 d. A positive reaction was
indicated by a clearing zone surrounding the growth of the organism. If no clearing zone was detected, the plates
were flooded with a 10% solution of trichloroacetic acid and re-examined.
The pH reactions of
isolates on agar plates were determined using YEM medium containing 0.25 mg/l
bromthymol blue.
Urease activity was
determined on urea agar slants (26) incubated for 7 d at 280C.
Citrate utilization was
determined on the solid medium of Koser (89).
Plates were spread with 108 cells, incubated at 300C
for 14 d, and examined for growth.
Penicillinase
(B-lactamase) was detected by the method of Foley and Perret (42), oxidase by
the method of Kovaks (90), and catalase by the method of Graham and Parker
(53).
Hydrogen sulfide
production was determined on agar slants [Hunter and Crecelius (72)]. Slants were inoculated and examined for H2S
after 14 d at 280C.
Nitrate reduction was
tested as described in the Manual of Methods for General Bacteriology (126), in
the same medium used by Graham and Parker (53) and in Difco nitrate broth.
For carbohydrate
utilization, the basal medium used (Bis) was that of Bishop et al.
(11) with different carbohydrates substituted for mannitol and 0.6 g/l KNO3
used as the nitrogen source. The medium
was solidified with purified agar (Difco).
All carbohydrates, with the exception of dextrin and starch, were filter
sterilized (0.4 um Nuclepore) before addition to gooled, molten, agar
medium. Dextrin and starch were added
to the medium before autoclaving at 1000C for 15 min. Each carbohydrate was added to a final
concentration of 1.0% (w/v). Inocula
were prepared by removing cells from YEM agar slants (with a cotton swab) and
suspending the cells to approximately 1X107 cells/ml in sterile
distilled water. A ten-fold dilution of
each cell suspension was added to the wells of a multiple inoculator plate
[Josey, et al. (81)] and inoculated onto the surface of carbohydrate
containing agar plates. Bishop’s agar
plates without carbohydrate served as controls. Duplicate plates of each carbohydrate were incubated at 280C
for 7 d and scored for growth.
D. Generation times in culture media and
sterile soil
Growth and pH responses
were determined in TY, Bis, YEM and PA (62) liquid media. All media were adjusted to pH 6.9 prior to autoclaving.
Fast-growing isolates were pre-grown (in the medium into which they would be
subsequently inoculated) for 3 d, while the slow-growers were pre-grown for 7 d. Inocula were added to an initial density of
106 cells/ml into 50 ml of the respective medium in 125 ml “side-arm”
Erlenmeyer flasks. Flasks were agitated
at 150 revolutions per minute (RPM) at 280C in a water-bath shaker. Cell growth was monitored using a
Klett-Summerson Photoelectric-Colorimeter (equipped with a # 66 red filter) and pH
determined after four days using an Orion Research (model 501) pH meter and a glass
combination-electrode.
Generation times in
sterile soil were determined using 10 g samples of air-dried Kula loam soil
(Typic Eutrandept, pH 6.5) in
70 ml screw-cap test tubes. Soil tubes
were autoclaved for 45 min
at 1210C, on two successive days, and inoculated with stationary
phase cultures of B. japonicum strains USDA 110 and 136, or fast-growing PRC
isolates USDA 193 and
205. Fifteen tubes of each organism
were inoculated to obtain initial cell numbers of about 5x105 cells/gm and a soil
moisture tension of about 60% of water-holding capacity. Cell growth was monitored by plate counts on
YEM agar using destructive samplings.
E. Intrinsic antibiotic resistances
Resistance to low levels
of antibiotics was determined using the method of Josey et al.
(81). Inocula were prepared as outlined
above for carbohydrates. Freshly
prepared, filter sterilized (0.4 um Nuclepore) solutions of antibiotics were
added to cooled, molten TY agar to give the following concentrations (ug/ml):
chloramphenicol 12.0, 25.0; kanamycin sulfate 10.0; naladixic acid 10.0; neomycin
2.5, polymyxin B sulfate 20.0, rifampicin 1.0, 6.0, streptomycin sulfate 2.5,
10.0; tetracycline-HCl 4.0; and vancomycin 1.5, 5.0. Controls consisted of TY agar plates without antibiotics. Isolates showing growth were scored as positive.
Duplicate plates of each antibiotic were incubated (in the dark) at 280C
for 7 d and scored for growth.
F. Enzyme Assays
6-phosphogluconate
dehydrogenase (EC 1.1.1.43) activity was determined in cultures grown for 72 h
at 260C in yeast extract-glucose medium (82). Cells were centrifuged at 6,000 X g for 10
min at 4oC and washed twice in 0.05 M sodium phosphate buffer, pH
7.4. Cell pellets were resuspended in
the same buffer, containing 2X10-4 M 2-mercaptoethanol (100) to a
final concentration of 1.0 g wet-weight cells/1.5 ml buffer and disrupted by
two passages through a French pressure cell at 15,000 pounds per square inch
(PSI) and cell debris removed by centrifugation at 14,000 X g for 30 min at 4oC. The clear supernate was stored at -200C
until use. The activity of NADP-linked
6-phosphogluconate dehydrogenase was measured by following the reduction of
NADP according to the method of Martinez-De Drets and Arias (101). Apparent endogenous enzyme activity was
subtracted from the results. Specific
activities were expressed in nanomoles of NADPH formed / min / mg of protein at
250C. Protein was determined
by the method of Lowry, et al. (98) using bovine serum albumin as
the standard.
B-galactosidase (EC
3.2.1.23) activity was determined in cultures grown for 3 d at 260C
in TY medium containing 0.5% (w/v) lactose.
Cells were centrifuged and washed as above and resuspended in 0.05 M
sodium phosphate buffer, pH 7.2, to a final concentration of 1.0 g wet-weight /
3.0 ml buffer and broken by passage through a French pressure cell at 15,000
PSI. Cell debris was removed as before
and supernatant fractions stored at 200C until use. Enzyme activity was measured by following
the appearance of a colored product (o-nitrophenol, ONP) at 420 nm (126). The incubation mixture (4.8 ml) contained:
2.7 ml enzyme reaction buffer [0.1 M sodium phosphate buffer (pH 7.0), 1X10-3
M MgSO4·7H20, 2X10-4 M MnSO4, and
0.05 M 2- mercaptoethanol]; 1.8 ml ONPG solution [0.1 M sodium phosphate buffer
(pH 7.0) and 1.3X10-2 M o-nitrophenyl-B-D-galactopyranoside (Sigma
Chemical Co. St. Louis, MO.)]; and 0.3 ml of cell-free extract. Enzyme assays were done at 370C
and the reactions stopped by the addition of 1.3 ml of reaction stop buffer
(8.0 M urea and 1.0 M Na2CO3, pH 12.0). Enzyme activities were expressed in
micromoles of ONP produced per min per mg of protein at 370C. Corrections were made for absorbance values
obtained in controls without substrate.
G. Ethanol utilization
The basal medium of
Bishop (Bis) (11), with or without added mannitol, and with 0.6 g/l KNO3
as the nitrogen source, was used in all ethanol experiments. Inocula for all studies were prepared by
gently washing the cells from YEM agar slants into 50 ml of Bis medium without
any carbon source. All cultures were “starved” by incubation
overnight at 28oC prior to inoculation. To determine whether strains could utilize ethanol as the sole
source of carbon and energy, starved cells were inoculated into 50 ml of Bis
(initial concentration approximately 106 cells/ml) supplemented
with 0.1, 0.25, 1.0, 2.0, 3.0, or 4.0% (v/v) ethanol. Cultures were examined after 7 d of incubation at 280C
and scored for growth. Bishop’s medium
without any carbon source served as control.
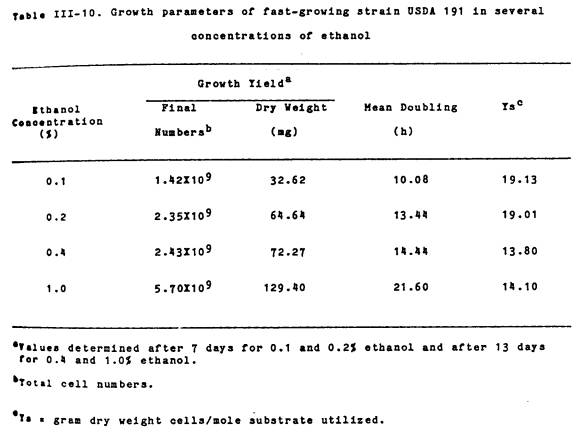
For growth yield and mean
generation time determinations when growing on ethanol, starved USDA 191 cells
were inoculated to approximately 106 cells/ml into 50 ml of Bis
supplemented with 0.1, 0.2, 0.4, or 1.0% ethanol and incubated at 28oC. Final, total cell numbers were determined
using a Petroff-Hausser counting chamber.
Mean generation times were calculated from the linear portion of growth
curves constructed by following absorbance at 600 nm. Growth yield values were determined after 7 d for 0.1 and 0.2%
ethanol and after 13 d for 0.4 and 1.0% ethanol. Substrate conversion values were determined by dividing the final
dry weight of cells in grams by the number of moles of substrate utilized. The disappearence of ethanol from the growth
medium was determined by gas chromatography using a Porpak Q column at 1850C
with N2 as the carrier gas.
For growth and survival
studies, 50 ml aliquots of Bis containing 0.2% ethanol were inoculated to about
1X106 cells/ml and incubated at 25oC. Samples from each culture flask were removed
at 0, 9, 14, and 18 days and viable cell numbers determined by plate counts on
YEM agar.
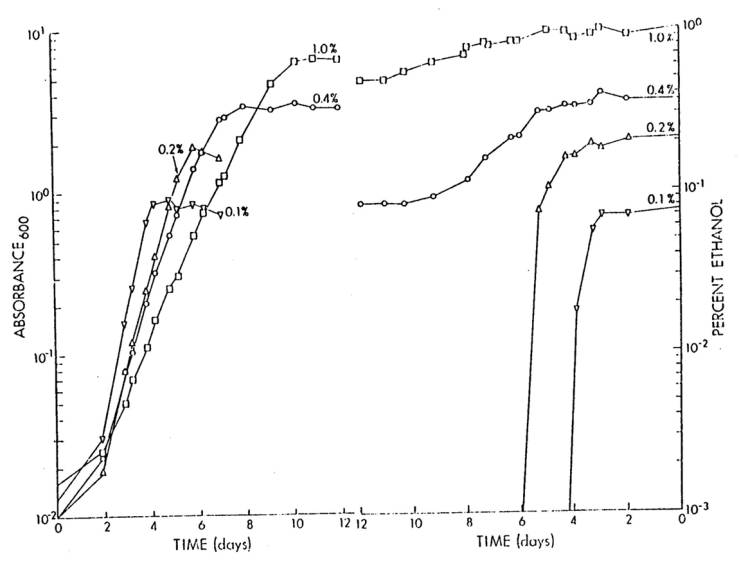
In experiments designed
to determine whether the lag phase of cells growing on limiting concentrations
of ethanol were affected by a subsequent addition of ethanol (at the same or at
a greater concentration), a starved USDA 191 culture was inoculated to a final
concentration of about 106 cells/ml into three, 500 ml side-arm
flasks each containing 100 ml of Bis medium.
Ethanol was added to two of the flasks to a final concentration of 0.1%
(a limiting concentration), while the remaining flask received 1.0%
ethanol. Cultures were incubated at 25oC
and growth was monitored using a Klett-Summerson Electric Colorimeter equipped
with a # 66 red filter. One of the two
flasks which initially contained 0.1% ethanol, received an additional 0.1%
ethanol at the beginning of each stationary phase of growth, while to the other
flask 1.0% ethanol was added only once.
The flask which originally contained 1.0% ethanol, received no further
ethanol additions.
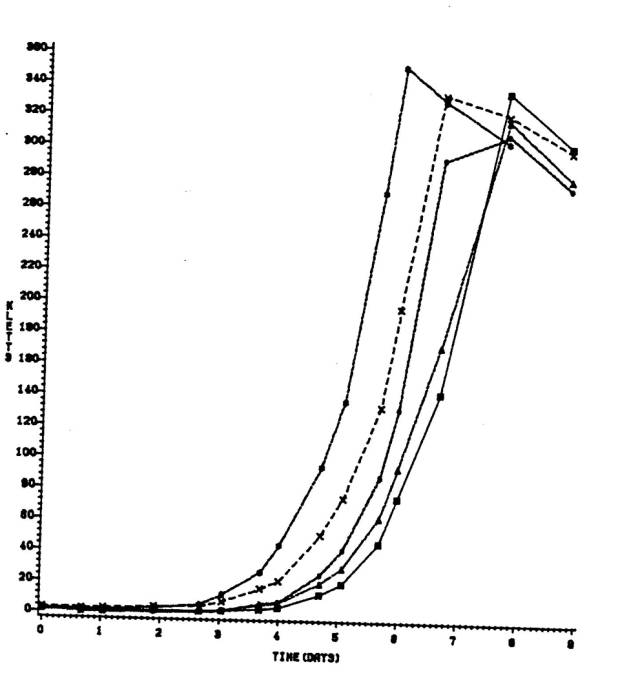
To determine if the
fast-growing soybean rhizobia were capable of showing diauxy when growing under
limiting concentrations of ethanol and mannitol, a starved culture of USDA 191
was inoculated into 75 ml of Bis supplemented with either 0.2% ethanol alone, 0.02%
mannitol alone, or 0.02% mannitol plus 0.2% ethanol. Flasks were inoculated, in triplicate, with approximately 106
cells/ml and two ml aliquots removed at different times to monitor growth
(absorbance at 600 nm), ethanol utilization by quantitative gas chromatography
(see above), and mannitol utilization by the
periodate-3-methyl-2-benzothiazolinone hydrazone method of Johnson and Sieburth
(75). For mannitol determinations,
periodate-digested samples and controls (samples without prior periodate digestion),
were analyzed in triplicate. All values
are the averages of three replicates.
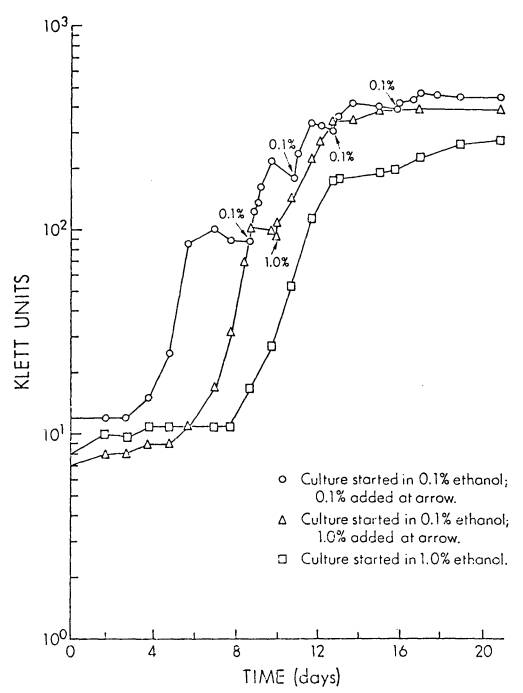
In experiments designed
to determine whether the inoculum size affected the length of the observed lag
phase, 125 ml side-arm flasks, containing 50 ml of Bis supplemented with 0.4%
ethanol, were inoculated, in triplicate, with USDA 191 to an initial cell
concentration of 1.6X105, 3.2X105, 6.4X105,
1.2X106, or 2.5X106 cells/ml. Initial cell numbers were determined by viable counts on YEM agar
and growth monitored spectrophotometrically using as Klett-Summerson
PhotoElectric Colorimeter equipped with a # 66 red filter.
Results
Morphological and cultural
characteristics. Both the fast-
and slow-growing soybean-rhizobia were Gram negative, non-sporeforming
rods. The fast-growing soybean isolates
tended to be slightly larger than the slow-growers, with their average
dimensions being 2 to 4 um by 0.5 to 1 um.
Cells from late log phase to stationary phase YEM cultures of fast-growers
tended to become enlarged and exhibited marked pleomorphism. Only a few cells (1-5%) of the fast-growing
rhizobia were motile in young (1-2 d) YEM cultures whereas the majority of
cells (80-90%) of the slow-growing rhizobia were motile. However, when growing
on the surface of moist B5 agar medium, a larger percentage (up to 25%) of the
fast-growing soybean rhizobia were found to be motile. On YEM agar plates containing brom thymol
blue, both fastand slow-growing rhizobia formed circular, convex, entire
colonies. After 6-7 d of growth, the
fast-growers formed colonies with sizes between 1.0 and 5.0 mm in diameter and
produced an acid-reaction, while the slow-growers had colony sizes of
approximately 0.5 to 1.0 mm in diameter and produced an
alkaline-reaction. Several of the
fast-growing soybean strains had a dry-crusty (calcified) appearance when grown
on YEM agar containing CaCO3.
In contrast to a large number of other fast-growing species of Rhizobium,
the fast-growing soybean rhizobia do not produce much extracellular
polysaccharides on YEM agar. Two of the isolates, USDA 191 and 192 produced “watery”
colonies. Upon repeated restreaking,
some of the fast-growers produced a second colony-type that was somewhat
smaller than colonies produced by the parent cultures. These colonial variants were not always
stable. Several of the variants that
did appear stable were isolated from seven cultures of fast-growers (USDA 191, 192, 193,
208, 214,
217, and 257) and tested for serological affinity (using strain-specific
fluorescent antibodies) and for effectiveness on soybeans (Glycine max)
cultivar Peking. They were found to be identical to the parental cultures.
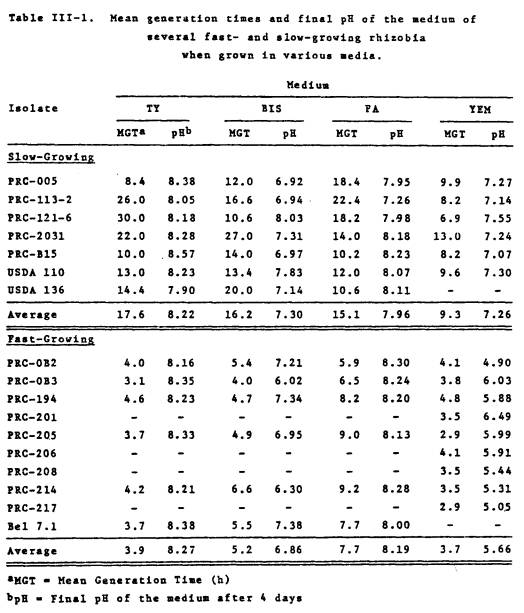
Generation times in culture
medium and sterile soil
All of the fast-growing
soybean rhizobia examined, had mean generation times much less than the
“typical” slowgrowers, in the four culture media examined (Table III-1). For the fast- and slow-growers, the most
rapid growth was attained in YEM medium, with an average generation time (MGT)
for the fast- and slow-growers of 3.6 and 9.3 h, respectively.
Results in Table III-1 also indicate that while TY medium is acceptable
for the growth of fast-growing rhizobia (MGT ranging from 3.1-4.2 h), most of
the slow-growers did not grow very well in this medium. The fast-growing PRC rhizobia acidified
only the YEM culture medium (average pH 5.6).
In the other three growth media, the pH either remained the same or
increased substantially. The
slow-growers on the other hand, raised the pH of all four culture media.
|
|
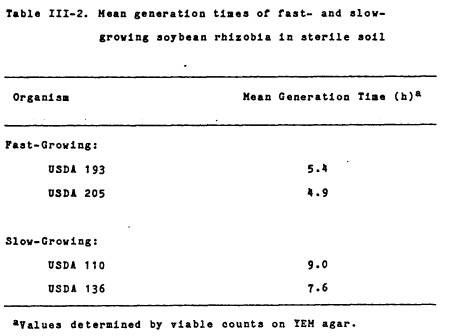
In sterile Kula loam soil,
the fast- and slow-growing soybean rhizobia had generation times (Table III-2) consistent
with those found in culture medium. The
two PRC isolates examined, USDA 193 and
205, had generation times only
slightly longer than those found in YEM culture medium, while the generation
time of B. japonicum strain USDA 110 was almost identical to that found
in YEM.
Growth responses to
pH and Nacl. Both
fast- and slow-growing soybean-rhizobia were examined for their ability to
grow in the presence of 2.0% NaCl and at pH extremes of 4.5 and 9.5.
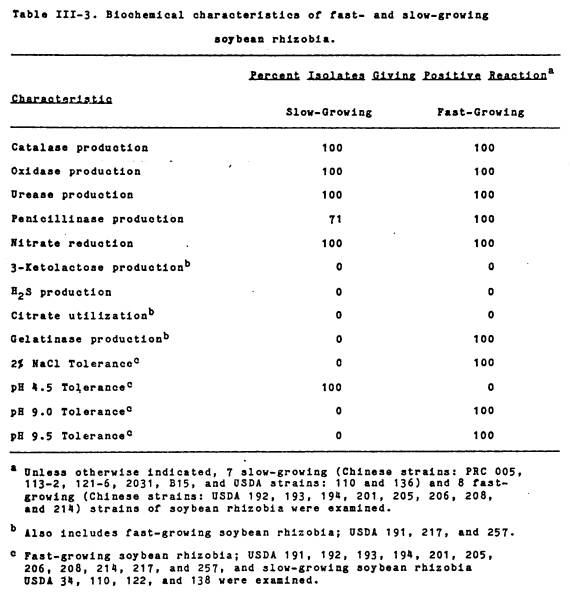
As is shown in Table III-3, the fast-growers were uniformly sensitive to the
low pH and tolerant of the high pH, while the inverse was found for the
slow-growers. The slow-growers grew
poorly at pH 9.0 and not at pH 9.5,
while the fast-growers grew well at pH 9.5. Conversely,
the slow-growers grew well at pH 4.5, while
the fast-growers did not grow at this pH.
The fast- and slow-growing rhizobia also differed with respect to their
NaCl tolerance. While all of the fast-growing isolates examined were able to
grow in the presence of the salt, there was variation among them in the extent
of growth. A few isolates (USDA 193,
194, 201 and 257) produced a confluent “lawn”
of growth on the medium, while others (USDA 191, 192, 205, 206, 208, 214, and
217) yielded individually tolerant colonies.
In contrast, none of the slow-growing B. japonicum grew in
2% NaCl.
Biochemical characteristics. The results shown in Table III-3 indicate
that both fast- and slow-growing rhizobia were catalase, oxidase, and urease
positive. All of the strains examined,
with the exception of the slow-growing PRC strains 2031 and B15, produced
penicillinase and all isolates reduced nitrate. None of the isolates produced 3-ketolactose
from lactose, hydrogen sulfide from Fe(NH4)2(SO4)2,
or utilized citrate as the sole source of carbon.
|
|
|
|
The two groups exhibited
a marked difference in gelatinase activity.
The fast-growing soybean-isolates produced a clearing zone on TY-gelatin
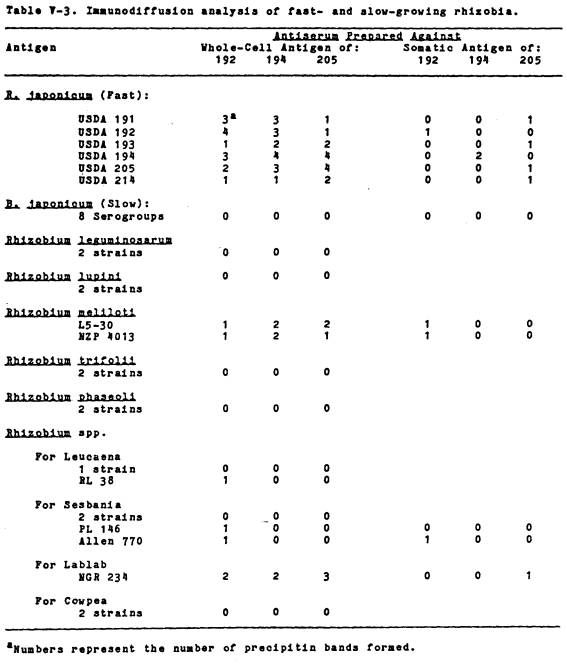
agar, while the slow-growers did not.
No gelatinase activity was detected in R. leguminosarum HI 5-0 and 92A3, R. phaseoli Bel 7.1, R. trifolii NZP 560, or Rhizobium
spp. USDA 3F4a4 and UMKL 19. However, a
Leucaena isolate, Tal 82, was positive.
The litmus milk reactions
of fast- and slow-growing rhizobia are shown in Table III-4. The slow-growing soybean rhizobia, with the
exception of PRC 121-6, exhibited no peptonization (serum zone formation) but
an alkaline pH change, while the fast-growers gave a variety of litmus milk
reactions. These included acid and
alkaline pH changes accompanied by peptonization, an alkaline pH change with a
lack of peptonization, and no pH change with peptonization. The fast-growing rhizobia tended to reduce
litmus, while the slow-growers did not. However, two fast-growing PRC isolates,
USDA 193 and 206, peptonized
litmus milk weakly, while one fast-growing isolate, USDA 208, failed to peptonize at
all. The other fast-growing rhizobia examined,
R. phaseoli Bel 7.1, R. phaseoli NZP 5097, R. leguminosarum 92A3, and 6015(pJB5JI), all
peptonized litmus milk, had neutral pH reactions, and variable litmus
reductions (data not shown).
Carbohydrate utilization. The results of carbohydrate utilization
presented in Table III-5 show that the fastgrowers utilized a greater variety
of carbohydrates than the slow-growers.
All of the organisms examined could utilize L-arabinose, D-fructose,
D-galactose, D-glucose, D-mannitol, D-mannose, L-rhamnose and D-xylose. The fast-growing soybean rhizobia, although
capable of utilizing arabinose, grew more slowly on this carbon source than the
slow-growers. Only the fast-growing
soybean-rhizobia utilized D-cellobiose, i-inositol, lactose, maltose,
raffinose, D-glucitol, sucrose, and D-trehalose. None of the
|
|
organisms examined could
utilize dextrin, galactitol, inulin, or starch as a sole source of carbon for
growth.
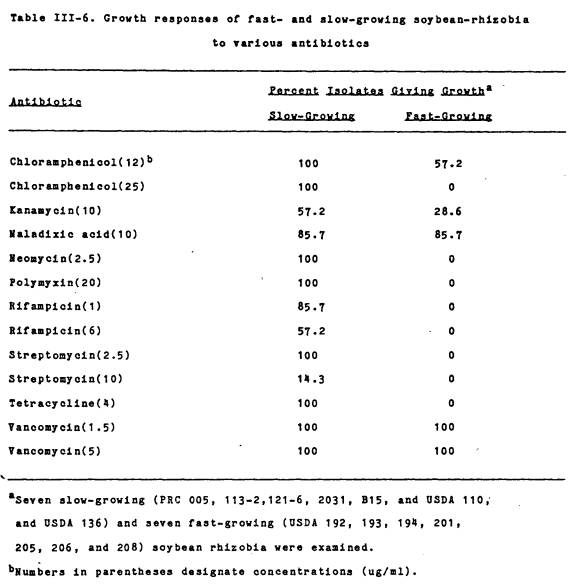
Antibiotic resistance patterns. The growth responses of fast- and
slow-growing soybean rhizobia to various anti-biotics are presented in Table
III-6. In general, the fastgrowing
rhizobia were sensitive to more antibiotics than the slow-growers. Two other fast-growing species examined, R.
leguminosarum 92A3 and R. phaseoli Bel 7.1, had growth
responses similar to the fast-growing soybean-rhizobia (data not shown).
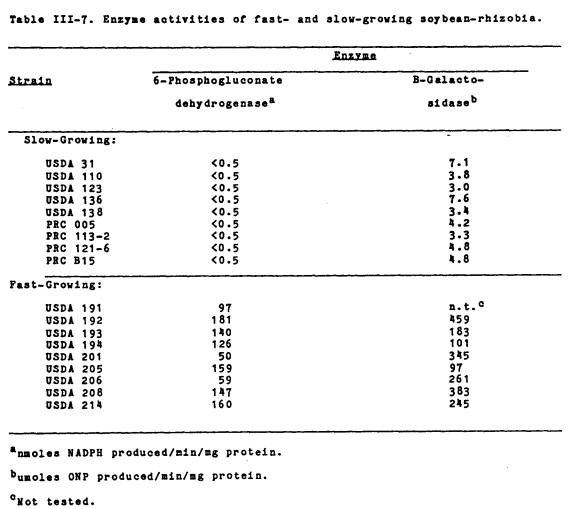
6-phosnhogluconate dehydrogenase (NADP-6PGD)
activity. The results, shown in
Table III-7, indicate that NADP-6PGD activity was only detected in the
fast-growing strains. The fast-growing
soybean rhizobia had NADP-6PGD activities ranging from 50 to 181 nmoles of NADP
reduced per min per mg of protein. No
activity was detected in the nine “typical” slow-growing B. japonicum
examined. The five other fast-growing Rhizobium
species tested (R. leguminosarum 92A3, R.
lupins NZP 2037, R. trifolii NZP 560, Rhizobium sp. Tal-82 and Rhizobium
sp. 3F4a4) had high specific activities for this enzyme, ranging from 26 to 156
nmoles NADP reduced per min per mg of protein.
B-Galactosidase activity. The results in Table III-7 indicate that
only the fast-growing strains had appreciable levels of B-galactosidase
activity. In general, they exhibited a
55-fold increase in enzyme activity over the slow-growers. The enzyme activity was comparable to that
found with the other fast-growing rhizobia (R. leguminosarum
92A3, R. phaseoli Bel 7.1, R. trifolii NZP 560, and Rhizobium sp. Tal-82) examined (data not shown).
|
|
|
|
|
|
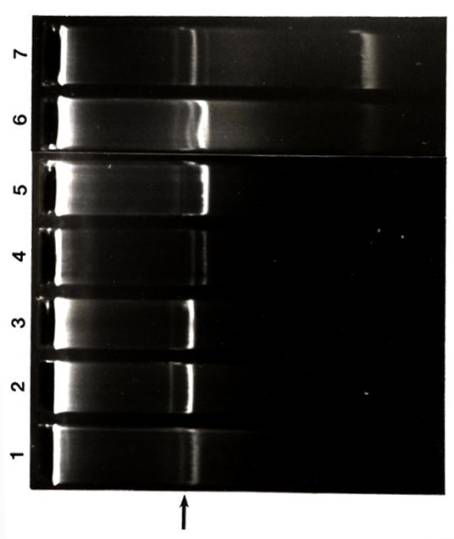
Growth in ethanol. Of the 30 strains from six species of Rhizobium
examined, only the fast-growing soybean rhizobia (USDA 191, 192, 193, 194, 201,
205, 206, 208, 214, and 217) were capable of substantial growth in ethanol
(Table III-8). One fast-growing
isolate, USDA 191, grew in ethanol up to a concentration of 3.0%. Of the slow-growing B. japonicum
tested (USDA 110, 123, 136, PRC 005, 113-2, 121-6, 2031, and B15) all, except
USDA 123, grew in 0.1% ethanol.
However, the slow-growers grew poorly and did not produce much
growth. The two slow-growing R. lupini,
CC814s and NZP 2021, were able to grow in 0.2% ethanol, while the fast-growing R.
lupini, NZP 2238 and SU 343, did not grow in ethanol at all. Two strains from each of the other species
of Rhizobium which were tested (R. legumiaosarum PRE and
92A3, R. meliloti L530 and NZP 4013, R. phaseoli
NZP 5253 and NZP 5260, and R. trifolii WU 95 and NZP 560) failed
to grow with ethanol as the sole source of carbon and energy.
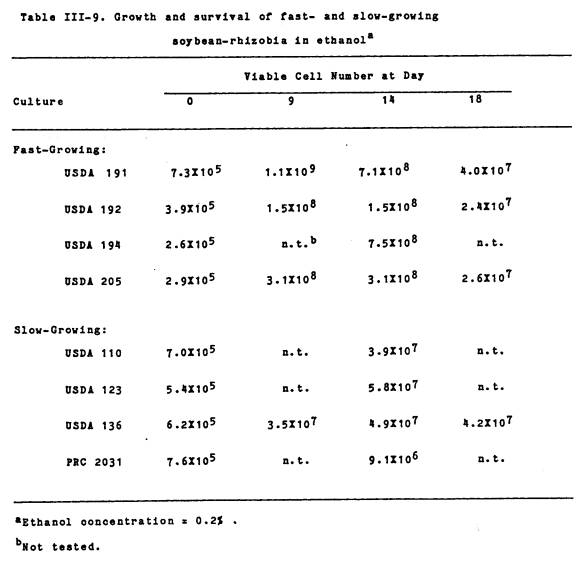
While the slow-growers
were capable of growing in 0.1% ethanol, final cell numbers never exceeded
about 4X107 cells/ml (Table III-9). On the other hand, all of the fast-growing
PRC rhizobia were capable of substantial growth in 0.2% ethanol, with final
cell numbers reaching between 108 109/ml. The slow-growing soybean-isolates survived
better in 0.2% ethanol than did the fast-growers. After 18 d of incubation, cell numbers for the fast-growers
decreased by factors of 10-100 from their 9 d values, while cell numbers for
the slow-growers remained about the same.
Fast-growing soybean
strains USDA 191 and 205 and slow-growing B. japonicum USDA 110,
were isolated from culture medium after growing in 1.0 and 0.25% ethanol,
respectively, and tested for nodulating ability. All of the isolates still were able to nodulate soybean cultivar
Peking (data not shown).
|
|
|
|
To determine whether
ethanol concentration affected the growth of the fast-growing soybean rhizobia,
USDA 191 cells were inoculated into Bishop’s medium containing 0.1, 0.2, 0.4,
or 1.0% ethanol. The results presented
in Table III-10 and Figure III-1 show that although ethanol can be used as a
sole source of carbon and energy by the the fast-growing soybean-rhizobia, it
may also be toxic to the cells at higher concentrations. Results presented in Table III-10 and Figure
III-1 show that growth is substrate limited with ethanol concentrations less
than 0.4%.
In previous experiments,
it was observed that the higher the ethanol concentration, the longer the lag
phase (data not shown). To determine
whether the length of the lag phase was correlated with the concentration of
ethanol and if preconditioning the cells in one ethanol concentration decreased
the lag phase in the same or a different concentration of ethanol, flasks
containing 0.1 or 1.0% ethanol were inoculated with USDA 191 (Figure
III-2). Cells in the two flasks
containing 0.1% ethanol showed logarithmic growth between 3-5 d, while
cells growing in 1.O% ethanol did not emerge from lag phase until about 8 d of
growth. Results in Figure III-2 show
that when 0.1% ethanol was added to cells previously growing on 0.1% ethanol,
there was a reduction in the length of the lag phase. Although, the addition of 1.0% ethanol reduced the growth rate of
cells previously growing in 0.1%, a lag phase following this addition was not
observed.
The results shown in
Figure III-3 indicate that the length of the lag phase is also related to the
intial inoculum size. With the
exception of one initial cell density, the greater the initial inoculum size,
the shorter the lag phase. Those flasks
receiving 2.5X106 cells/ml had a lag phase of about 2.7 d, while the flasks
receiving 1.6X105 cells/ml had a lag phase of about 5 d. Also, the
|
|
Figure III-1. Growth and ethanol utilization of USDA 191
in Bishop’s basal minimal medium with various concentrations of ethanol.
Figure: III-1A Growth; III-1B Ethanol utilization. Numbers on curves represent
initial ethanol concentrations in percent.
|
|
Figure III-2. Ethanol concentration step-up of USDA 191
while growing in two concentrations of ethanol. Cultures started in 0.1% ethanol, ○ and ∆, received
an additional 0.1% or 1.0% ethanol, respectively, at arrows. Culture started in 1.0% ethanol, □,
received no extra ethanol additions.
|
|
Figure III-3. Relationships between inoculum size and Lag
phase of USDA 191 when growing in 0.4% ethanol. Flasks were inoculated with: 1.6X105, □; 3.2X105,
○; 6.4X105, ∆; 1.3X106, X; or 2.6X106
*;cells/ml.
|
|
Figure III-4. Regression line of time to reach 15 Klett
units and Log number of inoculum size (cells/ml).
|
|
Figure III-5.
Growth and mannitol and ethanol utilization of USDA 191. Figure III-5A: Growth in 0.2% ethanol,
□; 0.02% mannitol, ○; and 0.02% mannitol plus 0.2% ethanol, ∆. Figure III-5B: Removal of ethanol from the
growth medium when growing in 0.2% ethanol, □ or in 0.2% ethanol plus
0.02% mannitol, ∆. Figure III-5C:
Removal of mannitol from the growth medium when growing in 0.02% mannitol,
○, or in 0.2% ethanol plus 0.02% mannitol, ∆.
|
|
graph of the time to
reach 15 Klett units vs Log inoculum size is linear (Figure III-4) with an R2
= 0.885.
To determine whether the
fast-growing soybean rhizobia would show diauxic growth when grown under
limiting concentrations of ethanol and mannitol, USDA 191 was inoculated into
0.2 % ethanol, 0.02% mannitol, and 0.2% ethanol plus 0.02% mannitol. Results in Figure III-5A show that when
cells were grown in limiting concentrations of mannitol alone, there was no
appreciable lag phase and cells reached stationary phase about 2 d after
inoculation. However, when cells were
grown in ethanol alone (also at a limiting concentration), the growth rate was slower
than that seen in mannitol, and it took about 5 d to reach stationary
phase. Cells grown under limiting
conditions of both mannitol and ethanol together, initially had a growth rate
similar to cells growing in mannitol alone, however, when mannitol became
limited, after about 2 d, the growth rate declined to about that seen for cells
growing on ethanol alone. The results
in Figure III-5B indicate that ethanol did not begin to disappear appreciably
from the growth medium untill after 3 d of growth and that ethanol utilization
was about the same for cells grown in ethanol alone or in ethanol plus mannitol.
Similarly, mannitol utilization was about the same for cells grown in mannitol
only, or cells grown in mannitol plus ethanol (Figure III-5C).
Discussion
Several diagnostic tests
were performed to compare fast-and slow-growing rhizobia that nodulate soybeans. The results indicate that the fast-growers
are quite different biochemically from the slow-growers. While the fast-growers
share host specificity with the slow-growers [both nodulate soybeans as well as
other hosts (84)], they appear more similar to other fast-growing Rhizobium
species in their microbiological, physiological and biochemical characteristics. Some characteristics, however, are shared
between the
two groups of organisms.
All of the fast- and
slow-growing soybean-rhizobia examined were typically positive for catalase,
oxidase, urease, penicillinase, and nitrate reductase. Both groups were unable to utilize citrate
as a sole source of carbon, produce H2S, or
produce 3-ketolactose. Graham and
Parker (53) also found that most Rhizobium japonicum strains they
tested possessed several of these attributes.
They reported that the production of H2S and
utilization of citrate was restricted to some isolates of R. meliloti. Bernaerts and De Ley (9) found that the
production of 3-ketolactose from lactose is limited to Agrobacterium species,
a genus closely related to Rhizobium.
Graham and Parker (53) found that the production of penicillinase seemed
to be restricted mainly to the slow-growing rhizobia, R. japonicum,
R. lupini and Rhizobium spp. of the “cowpea miscellany”
group. However, these authors indicated
that some of the R. leguminosarum strains also possess this attribute. Microbiologically, the fast and slow-growers
appear to be related in that they produce similar colony types (although they
do differ in size) and tend to produce morphological variants. However, as with other bacteria, the
production of morphological variants is not uncommon in Rhizobium
(58,134)
The PRC isolates were
truly fast-growing in sterile soil and in all of the laboratory culture media
examined. In most instances, when
growing in laboratory media, the mean generation times of the fast-growers were 2 to 3 times that of the
slow-growers. While the slow-growers
grew from 1.5 to 2 times faster in sterile soil than in media, growth rates of
the fast-growers in sterile soil was in the same range as that found in
laboratory media. It is interesting to
note that the fast-growing soybean isolates only reduced the pH of YEM medium,
while in all other media, the pH was raised.
This pH reduction-property can not only be due to the presence of
mannitol, since Bis also contained the same mannitol concentration. The reason(s) why Bis medium became alkaline
following growth of the fast-growers, is not understood. However, it is
important to remember, that for taxonomic considerations, determinations as to
whether rhizobia are alkaline producing or acid-producing should only be
ascertained in YEM medium.
The results of the pH and
salt tolerance tests place the fast-growing soybean-rhizobia with other
fast-growing rhizobia. Previous studies
of rhizobia (20,43,53),
in pure
culture and in soil, showed that the fast-growers were relatively more
alkali-tolerant and acid-sensitive, than the slow-growers. Also, Graham and Parker (53) showed that among
rhizobia, tolerance to 2% NaCl was restricted to the fast-growing R. meliloti. In addition, the responses of the
fast-growing soybean-rhizobia in litmus milk were more typical of fast-growing
Rhizobium, e.g. R. trifolii, R. leguminosarum, and R. meliloti (44,53).
Fast- and slow-growers
also appeared to be separated on the basis of their utilization of
carbohydrates and sensitivities to antibiotics. Graham and Parker (53) and
Fred et al. (44) showed that fast-growing rhizobia tend to use a
wider variety of carbohydrates than the slow-growers. This is clearly evident in Table III-5, with the fast-growers
using the majority of the carbohydrates examined and the slow-growers
relatively few. It should be noted that
the types of carbohydrates utilized also varies amongst the
soybean-rhizobia. As was pointed out by
Glenn and Dilworth (48), slow-growing rhizobia tend to lack both uptake systems
and catabolic enzymes for disaccharides.
My results are in agreement with these authors in that the disaccharides
cellobiose, lactose, maltose, trehalose, and sucrose and the trisaccharide
raffinose clearly separated the fast- from slow-growing rhizobia. The results in Table III-7 show that only
the fast-growing soybean rhizobia have appreciable B-galactosidase activities,
similar to that of the other fastgrowers examined (data not shown). These results agree with those of Glenn and
Dilworth (48) who found that among the rhizobia they examined, the inability of
slow-growers to utilize several disaccharides was due to the lack of disaccharide
uptake systems and the hydrolytic enzymes to cleave the disaccharides. The results in Table III-6 show that the
fast-growing soybean rhizobia were sensitive to more antibiotics than the
slow-growers. It should be noted that
the fast- and slow-growing rhizobia could not be placed into any resistance
groupings using the antibiotics and concentrations employed. Vincent (134) has indicated that while
strains within a species of Rhizobium show differential sensitivity to
antibiotics, the slow-growing species of Rhizobium are generally more
resistant than the fast-growing ones.
Consistent with the
division of rhizobia into fast- and slow-growing groups on the basis of
carbohydrates utilized, is their division based on the presence and absence of
enzymes of the pentose phosphate pathway.
As was pointed out by Martinez-De Drets and Arias (101,102), although
both fast- and slow-growing rhizobia have NAD-linked 6PGD activity, only the
fast-growers have NADP-linked 6PGD activity.
The results in Table III-7 demonstrate that the fast-growing
soybean-rhizobia have levels of NADP-6PGD activity comparable to those of the
other fast-growing rhizobia. These
results are consistent with those of Martinez-De Drets and Arias (100,101) and
Keele et al. (82) who found high levels of NADP-6PGD activity for
only fast-growing species of Rhizobium.
Although mannitol and
sucrose are the preferred carbon sources for fast-growing rhizobia (44) and
arabinose for the slow-growers (1), both groups are capable of utilizing many
different types of carbon compounds.
While it has been shown that in B. japonicum bacteroids
(the morphologic state of rhizobia within soybean root-nodules), aldehydes and alcohols
can support acetylene reduction and oxygen consumption (112), and that soybean
nodules contain acetaldehyde and ethanol (129), there have been no reports of
ethanol utilization by free-living rhizobia.
Results presented in this study indicate that both the fast- and
slow-growing soybean rhizobia are capable of utilizing ethanol as the sole
source of carbon and energy.
Although fast- and
slow-growing soybean rhizobia could not be separated on the basis of ethanol
utilization, there were some clear differences in the extent in which they used
ethanol. While the fast-growers
routinely grew to final cell densities of 1X108-1X109 on
0.2% ethanol, the slow-growers never reached densities greater than about 1X107
cells/ml on any concentration of ethanol.
It is apparent from results (Table III-10 and Figure III-1) showing
decreased growth rates in higher ethanol concentrations, that ethanol most
likely is toxic to the cells. The mean
generation time of USDA 191 increased nearly 2-fold, when the ethanol concentration
was raised from 0.1% to 1.0%. It is
interesting to note (Figure III-4) that whether the cells were originally
started in 1.0% ethanol or had their ethanol concentration raised from 0.1 to
1.0%, that their growth rates remained about the same. In the flask receiving multiple additions of
0.1% ethanol, growth rates also remained relatively constant following each
ethanol addition. The results presented
in Table
III-10 and Figure III-1 indicate that growth is substrate limited with ethanol
concentrations less than 0.4%. At ethanol
concentrations of 0.1-0.2%, final cell numbers and dry weight are proportional
to ethanol concentration. However, at
an ethanol concentration of 0.4% and greater, growth did not appear substrate
limited and growth yield values were not proportional to ethanol
concentration. Results in Figure III-1
show that the maximum rate of ethanol utilization (when 191 was growing in 0.1
and 0.2% ethanol) occurred after 3 to 4 d of growth. In 0.4 or 1.0% ethanol, the rate of utilization was considerably
less than that seen in lower concentrations.
Three of the fast-growing
soybean rhizobia (USDA 192, 194, and 205) were also examined for their ability
to use other low carbon-number compounds.
While all three of the isolates utilized acetate as a sole source of carbon,
only USDA 192 could use 1-propanol (data not presented). None of the organisms
examined could utilize methanol, formate, or 1-butanol as a sole source of
carbon (data not shown). While all of
the fast-growing soybean rhizobia utilized ethanol as a sole source of carbon,
I could not detect any NAD- or NADP- linked alcohol dehydrogenase (ADH)
activity in cellfree extracts of the organisms (data not shown). Rigaud and Trinchant (118) have demonstrated
a soluble alcohol dehydrogenase in R. meliloti. However, they did not examine whether cells
were capable of growth in ethanol.
Similarly, De Vries et al. (34) and Tajima and La Rue (129) demonstrated
the presence of an NAD-linked ADH in pea and soybean root nodule tissue,
respectively. The inabilty to detect
NAD or NADP-linked ADH in cell-free extracts of the fast-growing soybean
rhizobia may indicate that either the ADH of these organisms requires a
different coenzyme, or that the enzyme may not be present in cell-free extracts
(i.e., is membrane bound), or that the enzyme was present at undetectable
levels in the cells. Since malate
synthetase was detected in cell-free extracts of USDA 205 (data not shown), it
is assumed that they at least have some enzymes of the glyoxylate cycle. With this cycle present, the organisms have
a means for replenishing C-4 intermediates of the TCA cycle from acetyl CoA
derived from acetate, or ethanol. While
Johnson et al. (74) detected malate synthetase activity in
culture-grown B. japonicum cells, these authors failed to detect any
isocitrate lyase activity in these isolates.
However, isocitrate lyase was detected in culture-grown R. meliloti,
R. leguminosarum, R. trifolii, and R. phaseoli
cells (64) and in bacteroids from senescent soybean nodules (131). Interestingly, the organisms Johnson et
al. (74) examined could not utilize acetate as a sole source of
carbon. Thus, it is apparent that more
research is needed on the mechanism(s) by which fast- and slow-growing rhizobia
are able to utilize ethanol as a source of carbon.
In summary, while the
fast-growing soybean-rhizobia share symbiotic host-specificity with the typical slow-growers,
they appear more closely related, on a biochemical basis, to other fast-growing
species of Rhizobium. The typical
soybean-rhizobia are now classified as Bradyrhizobium japonicum
(79). Most of the evidence presented in
this thesis indicates that the newly isolated fast-growers do not fit into this
species. Therefore, the taxonomic
position of these new isolates must logically be in the new genus Rhizobium.
CHAPTER
IV
PLASMIDS
OF FAST-GROWING SOYBEAN RHIZOBIA: PROFILES
RESTRICTION
ENDONUCLEASE PATTERNS, AND ROLE IN MODULATION
Abstract
Several isolates from a
newly-described group of fast-growing acid-producing soybean rhizobia were
analyzed for plasmid content. All
contained from 1 to 4 plasmids with molecular weights of 100x106 or
larger. While most of the isolates
shared plasmids of similar size, the restriction endonuclease (BamHI, EcoRI
and HindIII) patterns of the plasmids from three of the isolates were
vastly different.
Growth in the presence of
acridine orange was effective in producing mutants cured of the largest plasmid
in one of the strains. These mutants had also lost the ability to form nodules
on soybeans. High-temperature curing of a smaller plasmid in another strain did
not lead to loss of nodulating ability or alteration of symbiotic effectiveness
on soybean cultivars.
The Rhizobium leguminosarum
host-range (Sym) plasmid, pJB5JI, is a 130 Mdal self-transmissible plasmid
which carries
the genes for host-specificity (peas) and some of the genes for
nitrogen-fixation. Plasmid pJB5JI was
transferred into different species of Rhizobium and the symbiotic
properties of the resulting transconjugants were examined. While,
transconjugants made with the fast-growing soybean-rhizobia were unable to
nodulate peas, they still retained the ability to nodulate soybeans. Transconjugants made with a Nod+
strain of R. trifolii, 0403, could nodulate clover (effectively)
and peas (ineffectively). However, the
transconjugants of a cured Nod- R. trifolii, #2, were
highly effective on peas, but still did not nodulate clover. After reisolation from pea and clover
nodules, #2 and 0403 transconjugants had a variety of symbiotic responses when
reinoculated onto peas and clover plants.
Evidence for successful transfer of the plasmid to all transconjugants
included: appearance of kanamycin resistance (due to TN5 in pJB5JI) in the
recipients, physical presence in the transconjugants of a new plasmid
corresponding to pJB5JI, and their ability to back-cross to a non-nodulating
strain of R. leguminosarum and make it infective. While plasmid pJB5JI is readily transferable
to many species of Rhizobium, its symbiotic performance varies depending
on the genetic background in which it resides.
Introduction
Recent advances in our
understanding of the genetics of the legume-Rhizobium symbiosis have,
for the most part, been achieved with the fast-growing species of Rhizobium:
R. leguminosarum (peas, lentils, and vetch) R. meliloti (alfalfa), R. phaseoli
(beans), and R. trifolii (clover) (10,17,18,35,76). This is in part due to the fact that symbiosis-related genes are
plasmid-borne in most of these rhizobia (10,62,65,66,108).
In 1967, Higashi (60) provided the first
suggestive evidence that plasmid genes are involved in nodulation and the
host-range specificity of Rhizobium trifolii. He found that R. trifolii and R. phaseoli lost their ability to
nodulate their respective hosts, following treatment with acridine orange. Acridine dyes were known (27) to cause elimination of
plasmids in other gram-negative bacteria and thus, it was assumed that the loss
of nodulating ability was due to the loss of resident plasmid(s). Similarly, Pariiskaya (111), and Zurkowski et
al. (141) demonstrated that growth in media containing acridine orange
resulted in the development of non-nodulating mutants. It was subsequently
shown (105,142) that Rhizobium cultures could be cured of their
nodulating ability by incubation at elevated temperatures (35-370C). However, it was not until the work of Nuti et al. (107), that physical evidence
for the presence of plasmids in Rhizobium was presented.
Rhizobia differ from most
other Gram-negative bacteria in that they possess very large plasmids
(megaplasmids). It is not unusual to
isolate Rhizobium plasmids with molecular weights (M.W.) between 100 and
300 megadaltons (Mdal) (107,114). Two R.
meliloti strains, T12 and T24, have been reported to carry plasmids of 190
and 460 Mdal (143). In all of the more
than 60 fast-growing Rhizobium strains thus far investigated, large
plasmids with molecular weights > 90 X106 have been reported
(33). Also, it is not unusual to
isolate more than one size class of plasmids from a single Rhizobium
strain. It is also interesting to note,
that the presence of plasmids with M.W.< 85 X106 have rarely been
reported in rhizobia. Thus, no resident
rhizobial plasmids are available as molecular cloning vehicles (33).
Little information is
available on the genetics of the slow-growing rhizobia, B. japonicum, R. lupini,
and Rhizobium sp. of the cowpea miscellany group. This is due partly to the inability to
demonstrate consistently plasmids in all infective strains. Furthermore, the location of
symbiosis-related genes in the slow-growing rhizobia is still unknown. Gross et al. (54) examined plasmids in
several isolates of B. japonicum (primarily from one serogroup). Although all isolates were obtained from soils with similar
characteristics and were predominantly from the same serogroup, 135, they
could be subdivided in 4 groups on the basis of plasmid number and size. However, these authors did not investigate the
location of symbiotic genes. Recently,
Masterson et al. (103)
reported that in the strains of B. japonicum that they
examined, the plasmids isolated did not contain genes that were hybridizable to
Klebsiella pneumoniae nitrogen fixation (Nif D and H) genes. Interestingly, these authors did detect
hybridization between NifDH genes and total cell DNA preparations in several
slowgrowing B. japonicum strains.
These results indicate that structural nitrogen fixation genes may not
be plasmid borne in these slow-growers.
Genetic studies involving
plasmid transfer in Rhizobium have almost exclusively been done with the
fast-growing species (8). Recently, Pilicinski and Schmidt (113) demonstrated
plasmid transfer within and between serologically distinct strains of B.
japonicum. However, the plasmids used in their study
were R68.45 and RP4 (originally from Pseudomonas), and not rhizobial
plasmids. Similarly, Kennedy et al.
(83) and Kuykendall (95) demonstrated the transfer of plasmids pRD1 and R68.45
from E. coli into several strains of slow-growing “cowpea”
rhizobia and B. japonicum, respectively. It should be noted however, that all of
these plasmids could not be transferred to all of the strains investigated.
The Rhizobium leguminosarum
plasmid, pJB5JI (pRL1JI::TN5), is the pea SYM (host-range) plasmid with a TN5
(kanamycin) insertion in the genes for medium bacteriocin production (76). It has been used by several investigators
(10,17,18,35,62,76,77) to transfer pea-nodulation genes into R. leguminosarum,
R. phaseoli, and R. trifolii
strains. Plasmid pJB5JI has a M.W.
of 130X106 and carries some of the nitrogen fixation (Nif) and
pea-nodulation genes (Nod). This
conjugal, self-transmissible plasmid also carries transfer function genes (Tra)
and is transferrable to a large number of Rhizobium strains at
frequencies of 10-1 to 10-3 (8). In addition, plasmid pJB5JI is in the same
compatibility group as other Rhizobium plasmids, and thus is maintained
intact in many recipients.
The present work
describes genetic investigations of the fast-growing soybean rhizobia. These investigations were undertaken to
determine: 1, whether the fast-growing PRC rhizobia contained large M.W.
plasmids; 2, whether there are structural relationships between plasmids from
different strains; 3, whether symbiosis-related genes are plasmidborne in these
isolates; and 4, whether the fast-growing soybean rhizobia were capable of
accepting, maintaining, and expressing plasmids from other fast-growing
rhizobia. In addition, this work
describes investigations into the stability, maintenance, and expression of a R.
leguminosarum Sym
plasmid in fast-growing soybean rhizobia as well as in R. trifolii
which have different genetic constitutions.
Materials
and Methods
A. Bacterial strains and maintenance of
cultures
The Rhizobium
cultures used in this study and their sources are listed in Table IV-1. All isolates (with the exception of USDA
110) were maintained on yeast extract mannitol, YEM, (132) agar slants containing
0.05% CaCO3. The
fast-growing soybean rhizobia chosen for this study fall into at least four
somatic serological groups (See Chapter V, This work).
B. Plasmid isolation and screening
Plasmid DNA was isolated
by a modification of the method described by Hirsch et al. (62). All buffers and labware were autoclaved before use. Cultures were grown for 48 h in 200 ml of PA medium (62),
supplemented with 0.20 g/1 of K2HP04. Bacteria were harvested by centrifugation at
10,000 X g and washed, two times, in TEN buffer (50 mm Tris, 20 mM Na2-EDTA,
and 50 mM NaCl, pH 8.0). The cell
pellet was resuspended in 16 ml of TEN buffer and lysozyme (Sigma Chemical Co.,
St. Louis) (5 mg/ml in sterile TEN) added to a final concentration of 0.55
mg/ml. The mixture was incubated at 37oC for 30 min and Pronase B
(CalBiochem Corp., California) (5 mg/ml in sterile TEN, pre-digested at 37oC for 1 h) and sodium
dodecyl sulfate (Sigma) [10% (w/v) in TEN] were added to final concentrations
of 0.5 mg/ml and 0.9% (v/v), respectively. The mixture was incubated at 37oC for 1 h or until lysis
was complete and the viscous lysate titrated to pH 12.4 by the addition of 3 M
NaOH with gentle, but thorough stirrinig (using the back-end of a plastic
knife). After incubation at room
|
|
temperature for 20 min,
the lysate was adjusted to pH 8.5 with 2 M Tris-HCl (Sigma) and transferred to
a 40 ml polycarbonate centrifuge tube.
Cold (4oC) 5 M NaCl was added to a final concentration of 1 M
and the contents of the tube mixed by gentle inversion. The mixture was incubated at 4oC
for 4 h and the SDS/NaCl precipitate removed by centrifugation at 10,000 X g
for 20 min at 4oC. The
supernatant was transferred to a 40 ml centrifuge tube and polyethylene glycol
(PEG) 6000 (Sigma) [50% (w/v) in TEN] was added to a final concentration of 10%
(v/v). The tube was mixed by inversion
and incubated overnight at 4oC and plasmid DNA collected by
centrifugation at 7,000 X g for 15 min at 4oC. The supernatant was gently decanted and the
pellet was allowed to dissolve in 0.5 ml of TEN buffer. Plasmid samples were stored at 4oC
for up to 1 week.
For preparative plasmid
extractions, all isolation steps were scaled up 10-fold and the plasmids were
further purified by CsCl-ethidium bromide (EtBr) equilibrium density gradient
centrifugation. Crude plasmid DNA, 4.5 ml, was added to 20 ml of CsCl-saturated
TEN buffer in a 40 ml centrifuge tube, the solution gently mixed by inversion
of the tube, and centrifuged at 10,000 X g for 20 min at 4oC. The resultant PEG/SDS precipitate was
removed from the upper portion of the solution and ethidium bromide (Sigma) (10
mg/ml in sterile distilled water) was added to a final concentration of 0.31
mg/ml. The refractive index of the
solution was adjusted to 1.3925+0.001 and transferred to 13 ml
nitrocellulose ultracentrifuge tubes (Beckman). The DNA preparation was centrifuged, 2 times, at 36,000 RPM and
170C for 48 hours, in
a Type 40 (Beckman) rotor (25). Ethidium
bromide and CsCl were removed from the plasmid preparation as is outlined by
Hirsch et al. (62) except that CsCl-saturated n-butanol was used
as the ethidium bromide extractant.
For plasmid screening,
samples were mixed with tracking dye [50%(w/v) glycerol, 0.125% (w/v)
bromphenol blue, and 50 mM Na2-EDTA, pH 8.0] and plasmids were
resolved by electrophoresis on horizontal 0.7% agarose (Biorad, Richmond, CA;
Standard Low) gels (18 by 13 by 0.6 em) at 60 mA for 6 h at 40C with
Tris-borate buffer (62). Gels were
stained for 20 min in Tris-borate buffer containing 5 ug/ml ethidium bromide
and bands were visualized by reflective, short-wave, UV light and photographed
using Kodak Pan-X film and a yellow (Hoya, K2) filter.
C. Restriction endonuclease analysis
For restriction enzyme
analysis, 60 ul of purified Plasmid DNA was digested with either BamHI, EcoRI,
or HindIII (Bethesda Research Labs, Inc.) for 1 h at 37oC according to the
suppliers directions. Following digestion, samples were heated at 65oC
for 3 min to inactivate endonucleases, and 15 ul of tracking dye was
added. Digested samples were
electrophoresed on 0.7% agarose gels at 50 mA for 16 h at 4o with
Tris-acetate buffer (65). Gels were
stained for 20 min in Tris-acetate containing 5 ug/ml ethidium bromide and
bands were visualized and photographed as before. For molecular weight estimations, the electrophoretic mobilities
of plasmid bands were compared to the reference strain R. leguminosarum
6015 (pJB5JI) (62,114) and the BamHI, EcoRI and HindIII
digests of lambda DNA (31).
D. Acridine orange and heat-curing of plasmid
DNA
Plasmid curing was done
by the acridine orange procedure of Zurkowski et al. (141) or by
the heat-treatment curing procedure of Zurkowski and Lorkiewicz (142). For acridine orange curing, overnight
tryptone yeast extract (TY) (62) cultures were diluted to approximately 104
cells/ml in YM medium (141) containing 1,3,7,15 or 20 ug/ml acridine orange
(Sigma Chemical Co.,St. Louis, Mo.).
Cultures were incubated at 25oC in the dark for 6 d. After incubation, cultures were diluted and
spread-plated onto YEM agar plates and the resulting colonies which formed were
streaked for purification two consecutive times on the same medium. For heat curing, overnight TY cultures were
inoculated to approximately 106 cells/ml into PA medium. Cultures were incubated at 37, 40, and 42oC
and transferred at weekly intervals.
After heat-treatment, cultures were spread-plated on YEM agar plates and
isolates purified as is outlined above.
E. Bacterial matings
Crosses were done
according to the membrane-filter method of Buchanan-Wollaston et al.
(22) from overnight TY cultures.
Aliquots from serial dilutions of the bacterial mixtures, harvested from
the membrane filters (with aid of a magnetic stir bar), were spread-plated onto
the appropriate selective media. To
select for R. japonicum and R. trifolii transconjugants
receiving plasmid pJB5JI (kanr), and against the auxotrophic (Phe,
Trp) donor 6015 (pJB5JI), the minimal Y-medium (3) supplemented with 50 or
100ug/ml kanamycin was used. In
back-crosses to the Nod- mutant, 6015 (Rifr,Strr),
the same medium was supplemented with kanamycin (50 ug/ml), rifampiein (20
ug/ml), streptomycin (100 ug/ml), phenylalanine (50 ug/ml), and tryptophan (50
ug/ml). This medium was called
Ysupp. In crosses between R.
trifolii (pJB5JI) and a Nod- USDA 205 (Strr,
Chlrr) mutant, the medium used was Y-medium supplemented with
kanamycin (100 ug/ ml), streptomycin (200 ug/ml), and chloramphenicol (10
ug/ml). All mixtures were also
spread-plated onto nonselective TY medium to obtain total viable counts.
F. Construction of antibiotic resistant mutants
Spontaneous antibiotic
resistant mutants were obtained by incubating cells, obtained from the surface
of YEM agar slants, in 100 ml of TY broth medium supplemented with various
concentrations and types of antibiotics.
G. Assessment of culture purity
Culture purity was
ascertained for all donors, recipients, and transconjugants in the following
manner: each culture was streaked two consecutive times on the appropriate
selective medium and isolated colonies transferred to TY liquid medium. After 2 d of growth, cultures were restreaked
on plates of selective media. Colonies
were transferred to slants of TY or TY supplemented with 50 ug/ml kanamyein,
as appropriate.
Each pure culture thus
obtained was further identified by immunofluorescence (123) or immunodiffusion
(132) using strain specific antiserum prepared according to Schmidt et al.
(123).
H. Plant infection assays
Nodulation tests for
soybeans and peas were performed in modified Leonard jar assemblies (96)
consisting of a 250 ml wide-mouth Erlenmeyer flask with a 13 mm hole in the
bottom. The flask was glued (silicone
seal) onto the cover (also having a 13 mm hole) of a 500 ml short, wide-mouth
screw-cap bottle, which served as the nutrient reservoir. An absorbant cotton wick was connected
between the two reservoirs to facilitate transfer of nutrient solution from the
lower reservoir into the plant growth vessel above. The growth vessel was filled with a Vermiculite:Perlite mixture
(3:1) and the nutrient reservoir with quarter-strength Hoagland's plant nutrient
solution (63). The modified flask
assembly was capped with aluminum foil and autoclaved before use. Clover nodulation was tested in screw-cap
tubes (2.5 by 20 cm) containing 25 ml of quarter-strength Hoagland's solution
(63) with 1.0% agar. The tubes were
slanted after sterilization. Peas (Pisum
sativa var Wisconsin Perfection) and soybean (Glycine max
vars Peking, Chippew a 64, and Lee) seeds
were surface sterilized by immersion in a 4.0% (w/v) calcium hypochlorite
solution for 20 min followed by exhaustive washings in sterile distilled
water. Three sterile seeds of the same
variety were aseptically transferred to each plant growth vessel and allowed to
germinate and grow until they reached the aluminum foil cap. Plants were selected for uniform size and
two of the three seedlings were aseptically removed from each vessel. Plants were inoculated with one ml aliquots
of 2-3 day-old YEM cultures and the vessels topped-off with approximately 2 cm
of sterilized Perlite and 2-3 cm of sterilized paraffin-coated sand (1.0 ml of
paraffin:chloroform (1:100) per 10 gm silica sand). Clover seeds (Trifolium repens var ladino) were surface
sterilized by immersion in a 2.0% solution of sodium hypochlorite for 10 min
followed by exhaustive washings in sterile distilled water. Seeds were
aseptically transferred to Petri dishes (150 by 25 cm) containing 1.0%
water-agar and germinated at 25oC in the dark. Seedlings were transferred
to Hoagland's-agar slants and inoculated with 0.1 ml of 2-3 day-old YEM liquid
cultures.
Plant vessels were
covered with aluminum foil to exclude light from the root zone and incubated
at 24+2oC in a plant growth chamber with an average light
intensity of 250 uEinsteins/m2/sec and a day length of 16 h. All plant tests were done in
triplicate. After four weeks of growth
(unless specified), plants were examined for the presence of nodules, for
color, and for nitrogenase activity by the acetylene reduction method (56).
I. Isolation and identification of nodule
occupants
For the recovery of
isolates from nodules, root sections (with attached nodules) were excised from
the plants and surface sterilized in a solution consisting of 75% ethanol and
8% H202. After
exhaustive washings in sterile distilled water, nodules were macerated in 10%
glycerol (peas and clover) or in distilled water (soybeans) and streaked onto
YEM agar containing 0.25 mg/1 brom thymol blue. Macerates were also spread onto
slides and nodule occupants examined by direct immunofluorescence using strain
specific antibodies (123).
Results
Plasmid profiles
All of the ten
fast-growing PRC isolates examined contained from 1 to 4 plasmids of
approximately 100 Mdal and greater.
Figure IV-1 shows the relative electrophoretic mobilities of plasmids
from USDA isolates 217, 208, 206, 193, 191, 201, 192, 214, 205 and 194 (Lanes:
A, D, E, F, G, H, J, K, L and M, respectively). The reference plasmids from R. leguminosarum
6015 (pJB5JI) appear in Fig. IV-1, Lanes C and I. All of the fast-growing PRC isolates with the exception of USDA
194 and USDA 191 shared a plasmid of similar size (approximately 200x106
Mdal). Isolates 217, 191, 201, 192,
205, and 194 each had 2 to 3
demonstratable plasmids (Fig. IV-1, Lanes A, G, H, J, L and M, respectively). One isolate, USDA 206, contained 4 plasmids
(Fig. IV-1, Lane E), while the other PRC isolates (Fig. IV-1, Lanes D, F and K)
contained one plasmid. Isolate USDA 191
(Fig. IV-1, Lane G) contained two plasmids, one of which exhibited the highest
electrophoretic mobility (lowest molecular weight) of all the PRC isolates
examined. No plasmids could be detected
in the slow-growing R. japonicum isolate USDA 110 (Fig. IV-1.
Lane N). However,
Figure. IV-1. Agarose gel electrophoresis of plasmid DNA
from fast- and slow-growing rhizobia.
Lanes A, D, E, F, G, H, J, K, L, and M fast-growing R. japonicum,
PRC isolates 217, 208, 206, OB3, 440, 201, OB2, 214, 205, and 194, respectively;
Lanes C and I, R. leguminosarum 6015(pJB5JI); Lanes B and N, slow-growing B. japonicum PRC
isolate 121-6 and USDA 110, respectively.
Numbers in margin refer to the size of the reference plasmids from
isolate 6015(PJB5JI) in Mdal (57,109).
|
|
another slow-growing R.
japonicum, isolate PRC 121-6 from China (Fig IV-1, Lane B), consistently
exhibited one plasmid with a molecular weight of about 100x106.
Restriction enzyme
analysis
Purified plasmid DNA from
USDA isolates 192, 194 and 205 [each a member of a separate serological group
(see Chapter 5)] was used for restriction enzyme analysis, with three
restriction endonucleases. Results in Figure IV-2 show that the EcoRI, HindIII
and BamHI (e, h and b, respectively), restriction patterns of plasmids
from the three isolates are vastly different from one another. For molecular weight comparisons, the
patterns of EcoRI, HindIII and BamHI digests of lambda DNA
are presented in Figure IV-2, Lanes IVe, h, and b, respectively. In all instances, restriction endonuclease
digestion gave rise to fragments with molecular weights less than 33 Mdal. The BamHI digest of plasmids from
USDA 205 (Fig. IV-2, Lane Ib) and the HindIII digest of plasmids from
PRC 194 (Fig. IV-2, Lane IIb) gave fragments with the highest molecular
weights. Although USDA isolates 205 and
194 (Fig. IV-2, Lanes I and II, respectively) contain plasmids of similar
number and size on Tris-acetate gels (Figure IV-2), their restriction digest
profiles clearly show them to be different.
These differences are most evident by the absence of restriction
fragments in several regions of the EcoRI, HindIII and BamHI
digests of plasmids from PRC 205 (Fig. IV-2, Lanes Ib, e, and h). Similarly, the restriction digest profile of
plasmids from USDA isolate 192 is different from either of the other two
isolates. It should also be noted that
after plasmid purification by CsCl centrifugation, one extra plasmid band of
approximately 33 Mdal became evident on Tris-acetate gels (Fig. IV-2, Lanes I
and II). This band was absent on
Tris-borate gels run with crude lysates (Fig. IV-1).
Figure. IV-2. Restriction
endonuclease digest of plasmids from fast-growing PRC R. japonicum. The undigested samples are given in lanes I
(PRC 205), II (PRC 194), and III (PRC OB2).
The three lanes following each number are digest patterns of e= Eco
R1, b= Bam H1, and h= HindIII restriction endonucleases. Intact lambda DNA and the respective
endonuclease digests are shown in lanes IV, e, h, and b, for comparison. Numbers in the margin refer to the size of
several lambda fragments in Mdal (28).
|
|
Curing of nodulation
plasmids
Three of the fast-growing
soybean isolates, USDA 192, 194, and 205, were used for acridine orange and
heat curing experiments. All three of
the isolates examined, failed to grow in acridine orange (AO) at concentrations
greater than 1 ug/ml. At an AO
concentration of 3 ug/ml or greater, viable cell numbers either decreased
slowly or remained the same from the initial inoculum level. Several isolates which were recovered from
growth-permissible (1 ug/ml) and restrictive ( >3 ug/ml) concentrations of
acridine orange were examined for their plasmid profiles and for their
nodulation responses on soybeans.
The plasmid pattern of
the “cured” mutants is shown in Table IV-2A. The results show that the AO
mutants of USDA 205 (205 1AO1, 1A03, and 1A05) are devoid of the largest
(approximately 200X106 Mdal) of the two plasmids. These mutants, isolated from cultures grown
in the presence of 1 ug/ml A0, and shown to be cured of their largest plasmid
failed to nodulate (Nod-) both soybean cultivars (Table IV-2B). However, all of the isolates recovered from
AO at 3 ug/ml and greater, effectively nodulated (Nod+,Fix+)
soybean cultivar Peking. Likewise, all of the USDA 194 and 192
isolates from 1 ug/ml AO retained their nodulating ability. The cured Nod- mutants of 205
were also used in blocking experiments to see whether they would inhibit
nodulation by Nod+ parental strain as well as other fast-growing R.
japonicum. There was
no significant suppression of nodulation even when the Nod- mutant
was inoculated 48 h before the addition of the parent strains.
Immunofluorescence
analysis of the somatic antigens of Nod- mutants indicated that the
mutants were serologically indentical with the parent strain. Immunodiffusion analysis using antisera
produced against somatic antigens as well as whole-cell antigens also indicated
that the nodulation deficient mutants were serologically identical to the
parent
|
|
(Table IV-2C). Both somatic and whole-cell antigens from
the mutants show patterns of identity with the parent strain.
Incubation of USDA 192
and 205 at elevated temperatures did not alter the plasmid profiles of the
recovered isolates. Organisms
recovered from growth medium incubated at 37 and 40oC for one week,
still nodulated both soybean cultivars and their plasmid profiles were
identical to their respective parents.
Similarly, incubation of USDA 194 at 37oC for 7 weeks or 42oC
for 1 week did not alter the plasmid profiles of the recovered mutants. However, one USDA 194 isolate, 194 1X40
(Table IV-2A), which was incubated at 40oC for 7 d, had a plasmid
profile different from the parent strain.
This isolate, was missing its lowest molecular weight plasmid band
(approximately 100X106 Mdal). However, it retained the ability to
nodulate soybeans. Immunofluorescence
and immunodiffusion analysis of its somatic and whole cell antigens showed that
it was serologically the same as the parental strain (Table IV-2C).
Frequency of kanamycin
resistance in fast-growing soybean rhizobia
Before examining the
ability of the fast-growing soybean isolates to receive plasmid pJB5JI (Tra+,
Nod+, Fix+, Kanr) from R.
leguminosarum strain 6015(pJB5JI), it was necessary to determine
if the isolates would grow on Y minimal medium supplemented with 50 ug/ml
kanamycin. Under conditions identical to those used in mating experiments, only
one, USDA 201, out of the seven isolates examined failed to grow in the
presence of 50 ug/ml kanamycin (Table IV-3).
The remaining six isolates, however, had natural kanamycin resistance
frequencies of 1.3x10-5 to 5x10-6. Three of the isolates, USDA 192, 201 and 205
(each a member of a different serological group) were also examined for their
ability to grow in the presence of higher concentrations of kanamycin. One isolate, USDA 192, grew at a kanamycin
concentration
|
|
of 100 ug/ml (frequency
of 3x10-6) while the other two isolates failed to grow at this
concentration. None of the three
isolates grew at a kanamycin concentration greater than 100 ug/ml (Table IV-3).
Plasmid transfer to
fast-growing PRC isolates
Fast-growing soybean
isolates, USDA 201 and 205, were examined for their ability to receive,
maintain, and express the pea-nodulating host-range plasmid, pJB5JI, from R.
leguminosarum 6015(pJB5JI). Transfer of the kanamycin marker to USDA 201
occurred at a frequency of 4.0X10-4 (Table IV-4) while transfer to
USDA 205 occurred at a frequency of 4.0X10-9. The low frequency of transfer to USDA 205
preeluded its use in further studies.
To confirm the presence
of pJB5JI in the USDA 201 transconjugants, two isolates 201-11 and 201-14, were
backcrossed to the Nod- R. leguminosarum strain,
6015. Transfer of pJB5JI from
transconjugants 201-11 and 201-14 to R. leguminosarum
6015 occurred at frequencies of 2.0X10-3 and 2.7X10-3,
respectively (Table IV-4). R.
leguminosarum 6015 transconjugants were picked from Ysupp plates
and purified as described previously.
In order to ascertain if
transconjugants received pJB5JI intact, or if the kanamycin resistance
phenotype was due to spontaneous mutation or transposition of TN5 from pJB5JI
(PRJ5JI::TN5) into the chromosome or the resident plasmids, plasmid DNA was
isolated from four USDA 201 and two 6015 transconjugants and visualized
following agarose gel electrophoresis.
Three classes of USDA 201 transconjugants could be detected by plasmid
profiles (Figure IV-3). The first class
consisted of those transconjugants, 201-8 and 201-13, which had plasmid
profiles similar to the parent strain USDA 201. Each of these isolates possessed two plasmids
|
|
p201a and p201b, of approximately 130 and 200 Mdal.,
respectively. The second class,
represented by transconjugant 201-11, had in addition to the two plasmids
present in USDA 201, a third smaller plasmid of approximately 100 Mdal. The third class of transconjugants,
represented by 201-14, appears to have lost its largest plasmid, but like
201-11, has gained an additional smaller plasmid. It should be noted that the smallest plasmid present in USDA 201
has an electrophoretic mobility similar to pJB5JI (Figure IV-3). The plasmid profiles of two back-crossed
USDA 6015 transconjugants, 6015-11-7 and 6015-14-8 along with their respective
parents are shown in Figure IV-4. Both
6015 transconjugants have received a plasmid band corresponding to plasmid
pJB5JI (130 Mdal) present in their respective parents (Figure IV-4).
Seven USDA 201
transconjugants, five 6015 transconjugants derived from back-crosses with
201-11, seven 6015 transconjugants derived from back-crosses with 201-14, and
whole mating mixtures from both 6015 crosses, along with USDA 201,
6015(pJB5JI), and 6015, were inoculated onto individual plants of peas and
soybeans (Glycine max var Peking). All seven of the USDA 201 transconjugants (Table IV-5)
effectively nodulated (Nod+, Fix+) soybeans, but failed
to nodulate peas. All of the 6015
back-crossed transconjugants examined effectively nodulated (Nod+,
Fix+) peas, whereas none of the transconjugants nodulated soybeans
(Table IV-5). Several 6015 transconjugants were reisolated from
nodules, and after purification, were reinoculated onto pea plants. All nodule-isolated transconjugants retained
the ability to nodulate peas. All
plants inoculated with whole mating mixtures also possessed effective
nodules. Plants inoculated with the
control mating mixtures of R. japonicum USDA 201 (parental) and R.
leguminosarum 6015 were not nodulated.
Two USDA 201
transconjugants, 201-11 and 201-14, were inoculated onto the commercial soybean
cultivars Chippewa and Lee to determine
Figure IV-3. Plasmid profiles of R. japonicum
USDA 201 and its transconjugants. Lane: 1, R. leguminosarum
6015(pJB5JI); 2, R. japonicum
201; 3, R. japnonicum 201-11(pJB5JI); 4, R. japonicum 201-8 kanr
; 5, R. japonicum 201-14(pJB5JI);
6, R. japonicum 201-13 kanr. Arrow indicates plasmid pJB5JI.
|
|
Figure IV-4. Plasmid profiles of R.
leguminosarum 6015 and its transconjugants. Lane 1: R. leguminosarum 6015(pJB5JI); 2, R.
japonicum USDA 201; 3, R. japonicum 201-11(pJB5JI); 4, R.
japonicum 201-14(pJB5JI); 5, R. leguminosarum 6015; 6, R.
leguminosarum 6015-14-8(pJB5JI); 7, R. leguminosarum
6015-11-7 (pJB5JI). Arrow indicates
plasmid pJB5JI.
|
|
|
|
whether the presence of
plasmid pJB5JI would now enable them to nodulate commercial varieties of
soybeans effectively. As is indicated
in Table IV-5, both transconjugants remained unable to produce an effective symbiosis
(Nod+, Fix-) with these cultivars.
Plasmid transfer to R.
trifolii
Rhizobium trifolii
isolates, 0403, and #2, were examined for their ability to receive, maintain,
and express the pea-nodulating, host-range plasmid, pJB5JI, from R. leguminosarum
6015(pJB5JI). R. trifolii
0403 is a “wildtype” clover-nodulating strain, while
R. trifolii #2 is a clover nodulatinn-deficient (Nod-)
strain. As before, both strains were
examined for natural resistance to kanamycin.
While strain #2 was unable to grow on Y medium with 50 ug/ml kanamycin,
strain 0403 was resistant to kanamycin at this concentration. However, at a kanamycin concentration
of 100 ug/ml, strain 0403 was resistant only at a frequency of 3.4X10-8
(Table IV-3).
Transfer of plasmid
pJB5JI from R. leguminosarum to R. trifolii 0403
and #2 occurred at frequencies of 2.1X10-4 and 4.4X10-4,
respectively (Table IV-4). Both of the
#2 transconjugants (2-33 and 2-54) and the one 0403 transeonJugant (403-33)
examined received a plasmid band corresponding in size to pJB5JI (Figure
IV-5). Interestingly, the “wild-type” parent strain 0403, was not found to
contain any plasmids despite repeated attempts at plasmid isolation and
visualization.
Six 0403 and #2
transconjugants, along with parental strains 0403, #2, and 6015(pJB5JI), were
inoculated onto peas and clover. All
six of the #2 transconjugants (Table IV-5) effectively nodulated (Nod+,
Fix+) peas, but still remained unable to nodulate clover. The six 0403 transconjugants examined
formed nodules on both peas and clover. While
Figure IV-5. Plasmid profiles of R.
trifolii strains #2 and 0403 and
their transconjugants. Lane: 1, R. leguminosarum
6015(pJB5JI); 2, R. trifolii
#2; R. trifolii 2-33(pJB5JI);
4, R. trifolii 2-54(pJB5JI);
5, R. trifolii 0403; 6,
R. trifolii 403-33(pJB5JI).
Arrow indicates plasmid pJB5JI.
|
|
clover nodules were
effective, all the nodules on peas, with the exception of one, were completely
ineffective. One of the three plants
inoculated with 0403 transconjugant 403-22, had one Fix+
nodule. All of the 0403 transconjugants
retained the ability to effectively nodulate clover (Nod+, Fix+)
(Table IV-5).
Two 0403 transconjugants
(403-42 and 403-33) and two #2 transconjugants (2-54 and .2-33) were
backcrossed to R. leguminosarum 6015. Plasmid pJB5JI
transferred from R. trifolii transconjugants 403-42, 403-33,
2-54, and 2-33 to R. leguminosarum strain 6015 at frequencies of
1X10-2, 1.4X10-2, 6X10-3, and 7.6X10-3,
respectively (Table IV-4).
Several R. leguminosarum
6015 transeonjugants derived from backcrosses to R. trifolii
transconjugants 403-42, 403-33, 2-54, and 2-33, were inoculated onto
individual plants of peas and clover.
All of the 6015 transeonjugants effectively nodulated (Nod+,
Fix+) peas, whereas none of the transconjugants nodulated (Nod-)
clover (Table IV-5).
To determine the
stability of plasmid pJB5JI in transconjugants after passage through plant
nodules, several 0403 and #2 transconjugants were reisolated from clover and
pea nodules and after purification and identification (using
immunofluorescence), were reinoculated onto individual plants of clover and
peas. The results shown in Table IV-6
indicate that the nodule-reisolated #2 and 0403 transconjugants produced a
variety of modulation and nitrogen fixation patterns on peas and clover
plants. After reisolation from pea and
clover nodules, some of the 0403 transconjugants (which originally formed an
ineffective symbiosis with peas and an effective symbiosis with clover) lost
their ability to modulate-peas, still produced an ineffective symbiosis, or
produced an effective symbiosis with peas.
All of the 0403 nodule reisolates (with the exception of 5 plants) still
retained the ability to effectively modulate clover. Interestingly, two of the #2 transconjugants gained the
|
|
ability to nodulate
clover after they were subsequently isolated from pea nodules. However, these transconjugants only produced
an ineffective symbiosis on clover.
Restoration of nodulating
ability
To determine whether pea
nodulating ability could be transferred to a fast-growing PRC isolate which has
lost its own nodulation plasmid, an acridine-orange-cured, nodulation-deficient
USDA 205 mutant [2051A03 (see above)] was used in crosses with R. leguminosarum
6015(pJB5JI) and R. trifolii 403-42 [0403 Chlr, Strr(pJB5JI)]. Despite repeated conjugation attempts, with
either donor, kanamycin resistant 2051A03 transconjugants could not be isolated
(data not shown).
Discussion
In this report, I have
demonstrated the presence of from 1 to 4 plasmids in ten fast-growing strains
of soybean-rhizobia. The recent
results of Masterson et al. (103) have indicated that several of
the fast-growing isolates contain plasmids which hybridize to the structural
nifK and D genes of Klebsiella pneumoniae. However, these authors did not show the location of nodulation
genes in these isolates. My results
suggest that in at least one of these isolates, a large plasmid may
also be the location of some of the nodulation genes. All of the acridine orange-generated,
nodulation-deficient mutants of USDA 205 had lost their largest plasmid. Zurkowski and Lorkiewicz (143) showed a
correlation between the loss of nodulation character and the elimination of a
plasmid in Rhizobium trifolii T12 and 24. Morrison, et al. (105) have
recently demonstrated that a Lablab purpureus isolate NGR234,
which can nodulate some legumes and the nonlegume Parasponia, could be
cured of its nodulating ability by treatment at elevated temperatures. In this isolate, the loss of nodulating
ability was found to result from the elimination of its largest resident
plasmid. In my study the heat-treated
USDA 194 mutant, missing a smaller plasmid, was still able to effectively
nodulate soybeans. Masterson et al.
(103) indicated that in USDA 194, the structural nif genes, were not
located on any of the plasmids.
Zurkowski and Lorkiewicz
(142) have shown that R. trifolii strains can easily lose their
nodulation plasmid at elevated temperatures.
The fast-growing soybean rhizobia that I examined did not lose their
nodulating ability at restrictive temperatures of 37oC for up to 7
weeks, or at 40 and 42oC for 7 d.
These isolates seem to be resilient to the heat-curing-treatments used
effectively for other species of Rhizobium. However, at levels of acridine orange that allowed slight growth,
one isolate was cured of its nodulating plasmid, while in bacteriostatic
concentrations, no curing of any plasmids could be demonstrated These results
are in agreement with those of Pariiskaya (111) who found that R. meliloti
L-1 was only significantly cured of modulating ability in subbaeteriostatic
levels of acridine orange. However,
Higashi (60)
and
Zurkowski et al. (141) cured different R. trifolii isolates of
their nodulating ability in bacteriocidal concentrations of acridine
orange.
The results of this study
indicate that plasmid pJB5JI, the pea-modulation, host-range plasmid from R. leguminosarum
6015(pJB5JI)
could be
transferred to the fastgrowing soybean rhizobia and R. trifolii
but was variably expressed in different genetic environments. When the plasmid was transferred to a
modulation-deficient isolate of R. trifolii, #2,
nitrogen-fixation and modulation genes were expressed, since the resulting
transconjugants effectively nodulated peas.
Initially, it was thought that plasmid pJB5JI did not restore the clover modulation deficiency present
in the #2 transconjugants. However,
after recovery from pea nodules, two of the transconjugants gained the ability
to nodulate clover. When pJB5JI was transferred to a
modulation-eom petent R. trifolii isolate, 0403, the resulting transconjugants were
capable of nodulating both clover and peas. However, peas (with the exception
of one plant) were only nodulated ineffectively (Nod+, Fix-). It should be noted however, that after
isolation from pea and clover nodules, there was a differential expression of
nodulation and nitrogen fixation genes among some of the 403
transconjugants. While one out of the
six transconjugants from clover nodules had lost the ability to nodulate peas
(most likely indicating a lost or deleted plasmid) three gained the ability to
effectively nodulate peas. While none
of the five 403 transconjugants reisolated from pea nodules lost the ability to
nodulate peas or clover, two of the reisolates could now nodulate peas
effectively. Thus, those plasmid-borne
genes required for an effective symbiosis, were differentially expressed in
some of the transconjugants after passage through host plants. The mechanism for this phenomenon is unknown.
Beynon et al. (10) noted that while some plasmids controlling
host-range specificity may be able to coexist (and behave one way) in culture,
they may not be able to do so within the nodules of a given host legume. That is, there may be some “physiological
incompatibility” or functional interference between the resident nitrogen
fixation and nodulation genes and those specified on plasmid pJB5JI. Likewise,
there may be functional interference between the clover nitrogen fixation genes
present in R. trifolii 0403, and the pea genes on pJB5JI, which
accounts for their lack of expression in 403 transconjugants. Passage through
the appropriate host plant may in some way modify this incompatability and
allow for the expression of both sets of genes.
Transfer of plasmid
pJB5JI to the fast-growing soybean isolate, USDA 201, did not bestow upon the
transconjugants the ability to nodulate peas. The presence of the
plasmid in the
transconjugants did not interfere with their ability to nodulate their own host
soybeans. However, in some transconjugants,
a loss of, or a deletion in, a resident plasmid was evident on agarose
gels. This did not, however, affect the
nodulating ability of the transconjugants, implying that the lost or deleted
fragments may not be involved in modulation.
While plasmid pJB5JI was transferable to fast-growing soybean strain
USDA 201, other isolates failed to accept the plasmid. One strain, USDA 205 was not able to receive
plasmid pJB5JI from R. leguminosarum 6015(pJB5JI) or R. trifolii
0403(pJB5JI) even when cured of its largest plasmid.
Plasmid pJB5JI has been
used by Brewin et al. (17, 18) and Hirsch et al.
(62) to transfer pea-nodulating ability to non-nodulating mutants of R. leguminosarum. Similarly, Beynon et al. (10) used
pJB5JI to transfer pea-nodulating ability to R. phaseoli and
Djordjevet et al. (35) used the same plasmid to transfer
pea-nodulating ability to R. trifolii. Even though the fast-growing USDA 201 transconJugants that
received pJB5JI do not themselves nodulate peas, they are, nevertheless,
capable of transferring this property to a non-nodulating strain of R.
leguminosarum, 6015.
This strain is a non-nodulating derivative of R. leguminosarum
strain 300 (77), similar to 6015(pJB5JI), but it is non-nodulating because of a
deletion in its largest Plasmid (62).
The smallest plasmid which appeared in 201-11 and 201-14
transconjugants, may be due to either
the co-transfer of another plasmid from R. leguminosarum 6015(pJB5JI), or a deletion in an incoming or resident
plasmid. Since USDA 201 contains a
plasmid which has a similar electrophoretic mobility as pJB5JI, deletions
in, or recombinational events between, this and other plasmids and the major
replicon are very probable. This may
explain why transconjugants 201-8 and 201-13 did not show any additional plasm
id bands despite their kanamycin resistance. However, the 6015 back-crosses,
6015-11-7 and 6015-14-8,
appear to have received pJB5JI from
201-11 and 201-14 intact as judged by the appearence of a new plasmid (Figure
IV-3) with an electrophoretic mobility similar to pJB5JI. Thus, the plasmid may have “recombined-out” of
201-11 and 201-14 during conjugation to R. leguminosarum 6015. As was indicated by Keyser et al.
(84), the fast-growing soybean rhizobia only form effective symbiosis with Glycine
Max var Peking and with Glycine
soja, but form ineffective nodules on most New World commercial
varieties. The results of this study
show that despite the presence of the extra nitrogen fixation genes (present in
pJB5JI), USDA 201 transconjugants still form ineffective nodules
on the two commercial cultivars of soybeans examined.
In conclusion, the
results of this study indicate that: 1, the loss of a large plasmid from the
fast-growing R. japonicum leads to loss of nodulating ability. These nodulation-deficient Rhizobium japonicum mutants
may prove useful in genetic manipulations of the soybean/Rhizobium
symbiotic system; 2, although the pea SYH plasmid, pJB5JI, is transferrable to
many different species of Rhizobium, the plasmid is differentially
expressed in different genetic backgrounds.
Despite the fact that the R. leguminosarum pea host-range
plasmid, pJB5JI, can effectively function in closely-related rhizobia, my
results suggest that it does not by itself carry all the genetic information
necessary for the nodulation of peas in a genetically dissimilar environment.
However, the plasmid is maintained intact in the different genetic backgrounds
despite its lack of expression; and 3, the host may play some role in
modifying the genetic make-up of the Rhizobium in the nodules, since
some of the nodule-reisolates had a different pattern of nodulation than the
parent cultures.
CHAPTER V
SEROLOGICAL
RELATEDNESS OF FAST-GROWING SOYBEAN RHIZOBIA TO
OTHER FAST- AND SLOW-GROWING
RHIZOBIA
ABSTRACT
Immunofluorescenee and immunodiffusion
reactions have been used extensively for the characterization of strains within
different species of Rhizobium.
The fast-growing soybean rhizobia from China (PRC) were examined to determine
their serological relatedness to each other, to the “typical” slow-growing B.
japonicum, and to other fast- and slow-growing rhizobia. Somatic
antigens of thirty-nine strains of Rhizobium were examined for
immunofluorescence cross-reactions with fluorescent antibodies (FAs) produced
against the somatic antigens of five fast-growing soybean-rhizobia.
Immunofluorescence (IF) analysis of seven of the fast-growing soybean strains
indicated that the isolates could be separated into at least three somatic
serogroups; 192, 194, and 205. One out
of the eleven typical slowgrowing B. japonicum (which were from
different serogroups) cross-reacted with one of the PRC FAs (194), while
members of the other serogroups failed to react with any of the FAs. The two
slow-growing cowpea rhizobia examined did not cross-react with any of the PRC
FAs. Of the 19 other fast-growing
rhizobia examined, one strain of Rhizobium sp. for Leucaena, Tal 82, and
three strains of Rhizobium spp. for Sesbania, Tal 1117, Allen 770, and
PL 146, fell within the 194 IF serogroup.
One strain of Rhizobium sp. for Lablab, NGR 234, cross-reacted
with all of the PRC FAs, however, iF cross-reactivity was greatest with FAs
prepared against USDA 192 and 205.
Somatic and whole cell antigens of 33 fast-
and slow-growing Rhizobium strains were tested for immunodiffusion (ID)
cross-reactions with antisera from three fast-growing soybean rhizobia. While
the six fast-growing soybean strains could be separated into at least three
somatic serogroups, they could not be grouped on the basis of whole-cell
reactions. The eight typical B. japonicum
(belonging to 8 different serogroups) and two slow-growing cowpea strains
failed to react with any of the whole- or somatic-cell antisera. While antigens from two strains of R.
leguminosarum, R. phaseoli, R. trifolii, and
R. lupini did not react with any of the antisera, several
fast-growing rhizobia clearly exhibited ID cross-reactivity. Antigens from two R. meliloti
strains and one strain of Rhizobium sp. for Lablab cross-reacted with
all three whole-cell antisera. Two out
of the four strains of Rhizobium sp. for Sesbania and one out of the two strains of Rhizobium
sp. for Leucaena cross-reacted with only USDA 192 whole-cell antiserum.
Results of this study indicate
that the fast-growing PRC soybean rhizobium are serologically related and can
be separated into at least three somatic serogroups using immunodiffusion or
immunofluorescence reactions. Also, that ID analysis using whole-cell antigens
indicates that the PRC isolates share at-least one antigen in common.
Introduction
Immunodiffusion, agglutination, and
fluorescent antibody techniques have been used widely to identify and characterize
strains of B. Japonicum grown in culture (36,23,37,49,125), present in nodules (15,24,37,38,104) and growing in soil (15). One major finding
of these studies is that the slow-growing soybean rhizobia exhibit a
high degree of serological specificity.
That is, strains of B. japonicum are serologically
heterogeneous (38,49). Another is that somatic antigens are more strain specific than
flagellar or internal antigens.
Koontz and Faber (88) and Wright (140) identified 6 somatic serogroups of B. japonicum using agglutination
reactions. However, Date and Decker (30) analyzed 28 strains of B. Japonicum and found 17 somatic serogroups on the basis of
cross-reactions and agglutination-adsorptions.
Skrdleta (124) found only four
somatic serogroups among the 40 strains he examined, but reported that 22 did not react with any of the 11 antisera
used.
While agglutination reaction was the first
serological method applied to the study of rhizobia, it lacks the resolving
power to distinguish between antigenically similar and closely related, but not
identical strains (38). Skrdleta (125), using
gel-immunodiffusions, divided the eleven B. Japonicum he examined
into two basic somatic serogroups. While he detected the same serogroups using
agglutinations, he found that immunodiffusions allowed him to show serological
relationships between strains which were not agglutinated by the same antisera.
Lastly, the fluorescent
antibody (FA) technique has been used by others (13,14,85,122,123) for the
direct examination of rhizobia in culture, nodules, and soils. The major advantages of FA over other
serological techniques is that only small amounts of antigen and antibody are
needed (16,121), the procedures are relatively rapid, and it is the only
technique which allows for the direct examination of rhizobia in situ (16).
This study describes a
serological investigation
of the
fast-growing soybean rhizobia for their degree of relatedness to each other,
to the “typical” slow-growing B. japonicum, and to other fast-
and slow-growing species of the root nodule bacteria.
Materials
and Methods
A. Bacterial strains, sources and maintenance
of cultures.
The strains used in this
study and their sources are listed in Table V-1. All cultures were maintained on YEM agar slants (132) (see
Chapter III for composition). Agar
slants used for the maintenance of fast-growing rhizobia contained 0.05% CaCO3. All cultures were incubated at 28oC.
B. Preparation of antigens.
For immunodiffusion (ID)
analysis, antigens were prepared from cells grown on the surface of B5 (45)
medium. Cells from the fast-growers
were harvested from 30 ml agar flats after 3 d of growth, while the slow-growers were harvested
after 7 d. Cells were resuspended in
2-3 ml of 0.85% saline (containing Thimerosal at a final concentration of
1:10,000) and stored at 4oC until use. For immunofluorescence (IF)
studies, antigens were prepared from cells grown in YEM liquid medium for 2 to
3 d. Somatic cell antigens for
injections and IF studies were prepared by placing cell suspensions in a
boiling water bath for 1 h.
|
|
C. Preparation of
antisera.
Boiled cell suspensions
were used to produce somatic-cell antisera according to the method of Schmidt et
al. (123). For the production of
whole-cell antisera, unboiled cell suspensions (whole-cell antigens) were
injected into the same rabbits previously used for the production of somatic
antisera. Rabbits were injected with one ml (0.5 ml in each hip) of an equal
mixture of antigen and Freund's complete adjuvant (Difco). After three weeks, two ml of the same
culture was injected intravenously without adjuvant. The rabbits were bled one week after the second injection.
D. Immunofluorescenee
Fluorescent antibodies
(FA) were prepared against the somatic cell components of USDA 192, 193, 194,
205, and 214. Antisera were conjugated
according to the procedures described previously (123).
Smears from pure cultures
and nodules were stained according to the method of Schmidt, et
al. (123). Gelatin-rhodamine
isothiocyanate conjugate (12) was used to suppress non-specific binding of
FA. Stained smears were examined with a
Zeiss standard 14 microscope equipped for epifluorescence and phase contrast.
Incident illumination was provided by an HBO-50 (OSRAM) mercury-vapor light
source with a fluorescein isothiocyanate (FITC) filter.
The cross-reactions of
FA-stained strains were assessed by subjective evaluation of the relative
intensity of fluorescence, from 0 to 4+, with 4+ being the highest degree of
fluorescence.
E. Immunodiffusion.
Immunodiffusion analyses
(ID) of whole and somatic cell antigens were performed according to the
procedures of Vincent (132). Cell
suspensions, containing 1010-1011 cells/ml were added to
the outer wells (6mm dia.) of Petri dishes containing 20 ml of immunodiffusion
agar [ 0.75% (w/v) Nobel agar (Difeo), 0.85% (w/v) NaCl, 0.01% (v/v)
Thimerosal, and 8X10-5 M Na2-EDTA, pH 6.5] and allowed to
prediffuse for 4 h before the addition of antiserum to the center well. Gels were incubated at room temperature for
3 to 4 d and stained with
amido black [0.1% amido black, 4.25X10-1 M acetic acid, 4.25X10-2
M sodium acetate, and 15% (w/v) glycerol]. Gels were destained with a 2.0%
(w/v) solution of acetic acid untill the background was clear and the resultant
precipitin bands which formed were recorded photographically.
Results
Immunofluorescence
Analysis
The results of the
immunofluorescence cross-reactions of 39 strains of fast- and slow-growing
rhizobia with 5 fluorescent antibodies are shown in Table V-2. The results presented indicate that the 7
fast-growing PRC soybean rhizobia can be separated into at least three somatic
serogroups. While the only member of
the 192 serogroup is USDA 192, the 194 serogroup consisted of USDA 194 and
201. The 205 serogroup consists of USDA
191, 193, 205, and 214.
Results of this study
indicate that with the fastgrowing PRC soybean-rhizobia, the intensities of
the immunofluorescence reactions were greatest when young (1-2 d old) YEM
cultures were used. If older cultures
were stained with FAs, iF reactions decreased in intensity and a smaller
percentage of cells reacted. In addition, the FA staining of the fast-growing
soybean strains differed from the other rhizobia examined in that cultures had
to be boiled, prior to FA staining (data not shown). Unboiled cells either failed to react or reacted poorly with the FAs.
Results of this study
also indicated while media other than YEM could be used for the growth of the
fast-growing soybean rhizobia, FAs did not stain cells grown in TY (62) or PPM
(see Chapter IV, this work). However,
FAs would stain cells grown in modified YEM medium where galactose had been
substituted for mannitol.
The five PRC FA's were
also used to determine if there were cross-reactions with other fast- and
slow-growing rhizobia (Table V-2). Of
the 11 slow-growing B. japonicum examined (comprising members of
11 different serotypes), only B. japonicum USDA 136 reacted with
one of the FAs (FA 194). Of the 19
other fast-growing rhizobia examined, one Rhizobium sp. for Leucaena,
strain Tal 82 and three Rhizobium sp. for Sesbania, strains Tal 1117,
Allen 770, and PL 146 fell within the 192 serogroup. One Rhizobium sp. for Lablab, NGR 234, cross-reacted with
all of the fast-growing PRC FA's.
However, the IF cross-reactivity of NGR 234 was greatest with FA's
prepared against USDA 192 and 205. Immunodiffusion analysis
The results of
immunodiffusion cross-reactions of 33 strains of Rhizobium with somatic
and whole cell antisera produced against USDA 192, 194, and 205 are summarized
in Table V-3. The results show that the
fast-growing soy bean-rhizobia could not be
|
|
separated into
serological groups based on their immunodiffusion reactions with the three PRC whole-cell antisera (see
Figure V-1 A for example). All of the
PRC strains examined shared at least one common heatlabile antigen. However, the fast-growing soybean strains
could be separated into at least three distinct serogroups based on ID
reactions with the three somatic antisera (see Figure V-1B for example). Generally speaking, the ID patterns obtained
with whole cell antisera were quite complex and involved at least four
discernable precipitin bands; while reactions with somatic antisera produced
1-2 bands.
While no precipitin bands
were detected in diffusion reactions between the 8 “typical” slow-growing
rhizobia and the three PRC whole or somatic cell antisera, several fastgrowing
rhizobia clearly exhibited ID cross-reactivity. Two R. meliloti strains, L5-30 and NZP 4013
produced 1-2 precipitin bands with whole-cell antisera from each of the
serogroups (192, 194, and 205), while the same isolates only produced one
precipitin band with somatic-cell antiserum from USDA 192. Although two Rhizobium spp. for
Sesbania, PL 146 and Allen 770, formed one precipitin band with whole-cell
antiserum against USDA 192, only Allen 770 reacted with 192 somatic-cell
antiserum. While strain NGR 234 of Rhizobium
sp. for Lablab (which also nodulates Leucaena and several other legumes) formed
precipitin bands with all three whole-cell antisera, it only gave one band with
somatic-cell antisera. No
immunodiffusion cross-reactivity was detected between the PRC antisera and any
other of fast- and slow-growing rhizobia examined.
|
|
Figure V-1. Serological analysis of several fastgrowing
soybean-rhizobia. Figure V-1 A. Immunodiffusion of whole-cell antigens of USDA 191, 192, 193, 194,
205, and 214 against antiserum
prepared from USDA 205 whole-cell antigens. Well: 1, USDA 192; 2, USDA 193;
3, USDA 191; 4, USDA 194; 5, USDA 205; 6, USDA 214. Center well contains wholecell antiserum against USDA 192. Figure V-1B.
Immunodiffusion of somatic antigens of USDA 191, 192, 193, 194, 205, and 214 against antiserum
prepared against the somatic antigens of USDA 192. Well: 1, USDA 192; 2, USDA 193; 3,
USDA 191; 4, USDA 194; 5, USDA 205; 6, USDA 214. Center well contains somatic-cell antiserum against USDA 192.
|
|
Discussion
Although the fast-growing soybean rhizobia
were isolated from geographically diverse regions of China, they were found to
be serologically related.
Immunodiffusion reactions with whole-cell antisera indicated that all of
the PRC isolates share at least one heat-labile agar diffusible antigen in
common. The heat-stable (somatic)
antigens analyzed by immunofluorescence or immunodiffusion, were useful in
separating the fast-growing soybean rhizobia into at least three serological
groups. Other workers (49,110,125,135)
have found that somatic antigens of fast- and slow-growing rhizobia are more
specific than flagellar or internal antigens and that these thermostable
antigens are suitable for separating strains within a species into distinct
serological groups.
The results of this study indicate that while
the fastgrowing soybean rhizobia share symbiotic host-specificity with the
typical slow-growing B. japonicum, they are not serologically
related. None of the strains from 8
serogroups of slow-growing soybean rhizobia examined, with the exception of B.
japonicum USDA 136 (CB 1809), cross-reacted with the five antisera
against the PRC strains. In addition,
antisera against the fast-growing PRC rhizobia did not react with two
slow-growing “cowpea” rhizobia or the one slowgrowing R. lupini isolates. It should be noted, that the cross-reaction
of USDA 136 with PRC 194 somatic antiserum was only detected by
immunofluorescence. No precipitin bands
could be detected in immunodiffusion reactions between USDA 136 and PRC 194
somatic antiserum. Pankhurst (110)
found that among the 62 fast- and 76 slow-growing Lotus rhizobia which he examined,
there was no sharing of somatic antigens between the two groups, when analyzed
by immunodiffusion reactions. The PRC
rhizobia showed some serological relatedness to some of the other fast-growing
rhizobia. Of the 19 fast-growing
rhizobia examined by immunodiffusion, 6 showed serological cross-reactivity
with the fast-growing PRC rhizobia. Out
of these six strains, the two R. meliloti strains and Rhizobium
sp. Allen 770, fell within the USDA 192 somatic serogroup, while the
fast-growing Lablab isolate, NGR 234 belonged to the PRC 205 somatic
serogroup. Similarly,
immunofluorescence analysis of 22 strains of fast-growing rhizobia revealed
that one Leucaena strain, Tal 82, and three Sesbania strains, Tal
1117, Allen 770, and PL 146 were members of the USDA 192 somatic
serogroup. On the other hand, the one
Lablab strain, NGR 234 examined by immunofluorescence, cross-reacted with all
five of the FA's used.
An interesting result of this study was the
finding that the two R. meliloti isolates cross-react with USDA
192 somatic antiserum. At present, all
available taxonomic and serologic evidence (see chapters 1, 2 and 5 this work)
indicate that R. meliloti are so unique among the
rhizobia, that they deserve separate species status. Vincent et al. (137)
have indicated that agglutination patterns of internal antigens placed strains
of R. meliloti with fast-growing rhizobia capable of nodulating
Lotus and Leucaena. Trinick (130) also
noted agglutination of R. meliloti by antisera prepared against a
strain of Leucaena rhizobia.
Another interesting finding was that while
the fast-growing rhizobia were capable of substantial growth in several
different laboratory media, they would not react with fluorescent antibodies
when the cells were grown in TY or PPM medium.
The phenomenon of antigenic variation due to cell growth conditions has
been reported by others. Vintikova et al. (139) indicated that
the nature of the growth medium affected the antigenic properties of R. meliloti. Dudman (38) also indicated that R. meliloti
antigens vary more than those of R. trifolii when grown in
different media. However, in this study
it was found that cells grown in TY medium would not even react with FA produced
from TY antigens (data not shown).
In summary: 1, although the fast-growing
soybean rhizobia were isolated from geographically different regions of China,
they are serologically related to each other and can be separated into at least
three distinct somatic serological groups; 2, immunodiffusion analysis of whole-cell
antigens revealed that all of the fast-growing PRC strains shared at least one
heat-labile antigen in common; and 3, somatic antigens were more specific than
whole cell antigens in separating these rhizobia into serogroups.
CHAPTER VI
GENERAL SUMMARY
The newly described fast-growing
soybean-rhizobia from China have provided a unique opportunity to study the biochemical,
physiological, and genetical relationships between fast- and slow-growing
rhizobia which have the ability to nodulate the same legume host. While soybeans represent a major
agricultural crop in the United States, China is thought to be the center of
origin and diversity of soybeans (and most likely soybean rhizobia).
The results of these investigations have
indicated that: 1, while the fast-growing soybean rhizobia share symbiotic
host-specificity with the typical slow-growers (they both nodulate the same
legume host), they appear more closely related, on a microbiological,
biochemical and physiological bases, to other fast-growing species of _Rhizobium.
Although the fast-growing soybean isolates share some biochemical
characteristics in common with other fast-growing rhizobia, there are some
biochemical properties (such as ethanol utilization and gelatinase activity)
which are unique to this group; 2, all of the fast-growing soybean isolates
which were examined fell within at least three somatic serogroups, based on
immunofluorescence reactions. While
immunodiffusion analysis of heat-labile whole-cell antigens indicated that all
the strains shared at least one heat-labile common antigen, immunodiffusions
performed with heat-stable somatic antigens were more specific and separated
the isolates into at least three somatic serogroups; 3, the taxonomic position
of the fast-growing soybean rhizobia must logically be in the new genus Rhizobium
and based on their symbiotic characteristics they may warrant separate species
status; 4, all the isolates of this group which were examined contain at least
one high molecular weight plasmid. In
one of the isolates the loss of a large resident plasmid led to the loss of
nodulating ability which indicates that nodulation genes might be plasmid borne
in this group of organisms; and 5, the pea host-range (Sym) plasmid, pJB5JI,
could be transferred to one of the fast-growing soybean isolates, but the
resulting transconjugants were unable to form nodules on peas. On the other hand, the transfer of pJB5JI to
two R. trifolii strains resulted in pea-nodulating
transconjugants indicating that the expression of the Sym plasmid depends on
the genetic background it resides in.
The fast-growing soybean rhizobia may provide
an effective tool for a better understanding of the genetics of the soybean-Rhizobium
symbiosis since they appear more amenable to genetic manipulations than the
typical slow-growing soybean symbionts.
LITERATURE CITED
1. Allen, E. E., and O. N. Allen. 1950.
Biochemical and symbiotic properties of the rhizobia. Bacteriol. Rev. 14:73-30.
2. Balassa,
G. 1963. Genetic transformation of Rhizobium: a review of the work
of R. Balassa. Bacteriol. Rev.
27:228-242.
3. Beringer,
J. E. 1974. R factor transfer in Rhizobium
leguminosarum. J. Gen. Microbiol.
84:188-198.
4. Beringer, J. E. and D. A. Hopewood. 1976.
Chromosomal recombination and mapping in Rhizobium leguminosarum. J. Gen Microbiol. 98:291-293.
5. Beringer,
J. E., A. W. B. Johnston, and B. Wells.
1977. The isolation of
conditional ineffective mutants of Rhizobium leguminosarum. J. Gen. Microbiol. 98:339-343.
6. Beringer, J. E., J. L. Beynon, A. V.
Buchanan-Wollaston, and A. W. B. Johnston. 1978. Transfer of drug
resistance transposon Tn5 to Rhizobium.
Nature 276:633-634.
7. Beringer,
J. E., S. A. Hoggan, and A. W. B. Johnston. 1978. Linkage mapping in Rhizobium
leguminosarum by means of R plasmid-mediated recombination. J. Gen. Microbiol. 104:201-207.
8. Beringer, J. E. 1980.
The development of Rhizobium genetics. J. Gen. Microbiol. 116:1-7.
9. Bernaerts, M. J., and J. De Ley. 1963.
A biochemical test for crown gall bacteria. Nature. 197:406-407.
10. Beynon, J. L., J. E. Beringer, and A. W. B.
Johnston. 1980. Plasmids and bost-range in Rhizobium leguminosarum
and Rhizobium phaseoli. J. Gen.
Microbiol. 120:421-429.
11. Bishop,
P. E., J. G. Guevara, J. A. Engelke, and H. J. Evans. 1976. Relation between glutamine synthetase
and nitrogenase activities in the symbiotic association between Rhizobium
jpoonicum and Glycine max.
Plant Physiol. 57:542-546.
12. Bohlool B. B., and E. L. Schmidt. 1968.
Nonspecific staining: its control in immunofluorescence examination of
soil. Science. 162:1012-1014.
13. Bohlool,
B. B., and E. L. Schmidt.
1970. Immunofluoreseent
detection of Rhizobium japonicum in soils. Soil Sci. 110:229-236.
14. Bohlool,
B. B., and E. L. Schmidt.
1973. A fluorescent antibody
technique for determination of growth rates of bacteria in soil. Bull. Eco. Res. Comm. (Stockholm). 17:336-338.
15. Bohlool,
B. B., and E. L. Schmidt.
1973. Persistance and
competition aspects of Rhizobium japonicum observed in soil by
immunofluorescence microscopy. Soil
Sci. Soc. Am. Proc. 37:561-564.
16. Bohlool,
B. B., and E. L. Schmidt.
1980. The immunofluorescence
approach in microbial ecology, p.203-241. In M. Alexander (ed.),
Advances in Microbial Ecology, Vol. 4. Plenum Press, New York.
17. Brewin,
K. J., J. E. Beringer, and A. W. B. Johnston. 1980. Plasmid-mediated
transfer of host-range specificity between two strains of Rhizobium leguminosarum. J. Gen. Microbiol. 120:413-420.
18. Brewin,
K. J., J. E. Beringer, A. V. Buchanan-Wollaston, A. W. B. Johnston, and P. R.
Hirsch. 1980. Transfer of symbiotic genes with
bacteriocinogenic plasmids in Rhizobium leguminosarum. J. Gen. Microbiol. 116:261-270.
19. Brewin,
K. J., T. M. Dejong, D. A. Phillips, and A. W. B. Johnston. 1980.
Co-transfer of determinants for hydrogenase activity and nodulation
ability in Rhizobium leguminosarum. Nature 288:77-79.
20. Bryan,
U. C. 1923. Effects of acid soils on noduleforming
bacteria. Soil Sci. 15:37-40.
21. Buchanan-Wollaston, V. 1979.
Generalized transduction in Rhizobium leguminosarum. J. Gen. Microbiol. 112:135142.
22. Buchanan-Wollaston,
A. V., J. E. Beringer, N. J. Brewin, P. R. Hirsch, and A. W. B. Johnston. 1980.
Isolation of symbiotically defective mutants of Rhizobium leguminosarum
by insertion of the transposon Tn5 into a transmissible plasmid. Mol. Gen. Genet. 178:185-190.
23. Bushnell,
0. A., and W. B. Sarles. 1939. Investigations upon the antigenic
relationships among the root-nodule bacteria of the soybean, cowpea, and lupine
cross-inoculation groups. J.
Bacteriol. 38:401-410.
24. Caldwell,
B. E. and G. Vest. 1968. Nodulation interactions between soybean
genotypes and serogroups of Rhizobium japonicum. Crop Sci. 8:680-688.
25. Casse,
F., C. Boucher, J. S. Julliot, M. Michel, and J. Denarie. 1979. Identification and characterization of large
plasmids in Rhizobium meliloti using agarose gel
electrophoresis. J. Gen. Microbiol.
113:229-242.
26. Christensen,
W. B. 1946. Urea decomposition as a means of
differentiating Proteus and paracolon organisms from each other and from
Salmonella and Shigella types.
J. Bacteriol. 52:461-466.
27. Cloves, R. C., E. E. M. Moody, and R. H.
Pritchard. 1965.
The elimination of extrachromosomal elements in thy mineless strains of Escherichia
coli K12. Genet. Res., Camb. 6:147.
28.
Craig,
A. S., R. M. Greenwood, and K. I. Williamson. 1973. Ultrastructural inclusions of rhizobial bacteroid of Lotus
nodules and their taxonomic significance.
Arch. Mickrobiol. 89:23-32.
29.
Crow,
V. L., B. D. W. Jarvis, and R.M. Greenwood. 1981. Deoxyribonucleic acid homologies among acid-producing
strains of Rhizobium. Int. J.
Syst. Bacteriol. 31:152-172.
30. Date, R. A., and A. M. Decker. 1965.
Minimal antigenic constitution of 28 strains
of Rhizobium japonicum.
Can. J. Microbiol. 11:1-8.
31. Davis, R. W., D. Botstein, and J. R. Roth. 1980.
A manual for genetic engineering; Advanced bacterial genetics, p. 236. Cold
Spring Harbor Laboratory, Cold Spring Harbor, New York.
32. De
Ley, J., and A. Rassel. 1965. DNA base composition, flagellation, and
taxonomy of the genus Rhizobium.
J. Gen. Microbiol. 41:85-91.
33. Denarie,
J., P. Boistard, F. Casse-Delbert, A. G. Atherly, J. O. Berry, and P. Russell. 1981.
Indigenous plasmids of Rhizobium, p. 225-246. In K. L. Giles and A. G. Atherly
(ed.), Biology of the Rhizobiaceae, Suppl. 13. Academic Press, New York.
34. De
Vries, G. E., P. IN ‘T Veld, and J. W. Kijne. 1980. Production of organic acids in Pisum
sativum root nodules as a result of oxygen stress. Plant Sci. Lett. 20:115-123.
35. Djordjevic,
M. A., W. Zurkowski, and B. G. Rolfe.
1982. Plasmids and stability of
symbiotic properties of Rhizobium trifolii. J. Bacteriol. 151:560-568.
36. Dudman,
W. F. 1964. Immunodiffusion analysis of the
extracellular soluble antigens of two strains of Rhizobium meliloti. J. Bacteriol. 88:782-794.
37. Dudman,
W. F. 1971. Antigenic analysis of Rhizobium japonicum
by immunodiffusion. Appl. Microbiol.
21:973-985.
38. Dudman,
W. F. 1977. Serological methods and their application to
dinitrogen-fixing organisms, p. 487-508.
In R. W. F. Hardy and A. H. Gibson (ed.), A Treatise On
Dinitrogen Fixation, Section IV. John
Wiley and Sons, New York.
39. Dunican,
M. J., and A. B. Tierney.
1974. Genetic transfer of
nitrogen fixation, from Rhizobium trifolii to Klebsiella
aerogenes. Biochem. Biophys. Res.
Comm. 57:62-72.
40. Elkan,
G. H. 1969. Deoxyribonucleic acid base compostion of
isolates of Rhizobium japonicum.
Can. J. Microbiol. 15:490-493.
41. Elkan,
G. H. 1981. The taxonomy of the Rhizobiaceae, p.
1-14. In K. L. Giles and A. A., Atherly (ed.), Biology of the Rhizobiaceae,
Suppl. 13. Academic Press, New York.
42. Foley,
J. M., and C. J. Perret. 1962. Screening bacterial colonies for
penicillinase production. Nature
195:287-288.
43. Fred,
E. B., and A. Davenport. 1918. Influence of reaction on
nitrogen-assimilating bacteria. J. Ag.
Res. 14:317-336.
44. Fred,
E. B., I. L. Baldwin, and E. McCoy.
1932. Root nodule bacteria and
leguminous plants. University of
Wisconsin Studies in Science, Number 5, Madison.
45. Gamborg, 0. L. 1975. Callus
and cell cultures, p. 1-9. In 0. L. Gamborg and L. R.
Wetter (ed.), Plant Tissue Culture Methods. National Research Council of
Canada, Saskatchewan.
46. Gibbins,
L. N. 1967. The preparation of antigens of Rhizobium
meliloti by ultrasonic disruption: an anomaly. Can J. Microbiol. 13:1375-1379.
47. Gibbins,
A. M., and K. F. Gregory. 1972.
Relatedness among Rhizobium and Agrobacterium species
determined by three methods of nucleic acid hybridization. J. Bacteriol. 111:129-141.
48. Glenn,
A. R., and M. J. Dilworth.
1981. The uptake and hydrolysis
of disaccharides by fast- and slow-growing species of Rhizobium. Arch. Microbiol. 129:233-239.
49. Graham,
P. H. 1963. Antigenic affinities of the rootnodule
bacteria of legumes. Antonie van
Leeuwenhoek, J. Microbiol. Serol. 29:281-291.
50. Graham,
P. H. 1964. The application of computer
techniques to the taxonomy of the root-nodule bacteria of legumes. J. Gen. Microbiol. 35:511-517.
51. Graham,
P. H. 1971. Serological studies with Agrobacterium
radiobacter, A. tumefaciens, and Rhizobium strains. Arch. Mikrobiol. 78:70-75.
52. Graham,
P. H., and M. O’Brien. 1968. Composition of lipopolysaccharides from Rhizobium
and Agrobacterium. Antonie van
Leeuwenhoek 34:326-330.
53. Graham,
P. H., and C. A. Parker. 1964. Diagnostic features in the characterization
of the root-nodule bacteria of legumes.
Plant Soil 20:383-396.
54. Gross,
D. C., A. K. Klucas, and R. V. Klucas.
1979. Plasmids, biological
properties and efficacy of nitrogen fixation in Rhizobium japonicum
strains indigenous to alkaline soils. J.
Gen. Microbiol. 114:257-266.
55. Hadley,
H. H., and T. Hymowitz. 1973. Chapter 3. Speciation and cytogenetics, p.
97-116. In B. E. Caldwell (ed.),
Soybeans: Improvement, Production, and Uses.
Am. Soc. Agron., Madison, Wisconsin.
56. Hardy,
R. W. F., R. D. Holsten, E. K. Jackson, and R. C. Burns. 1968.
The C2H2-C2H4 assay for
nitrogen fixation; laboratory and field evaluation. Pl. Physiol. 43:1185-1207.
57. Heberlein,
G. T., J. De Ley, and R. Tijtgat.
1967. Deoxyribonucleic acid homology and taxonomy of Agrobacterium,
Rhizobium, and Chromobaeterium.
J. Bacteriol. 94:116-124.
58.
Herridge,
D. F., and R. J. Roughley. 1975.
Variation in colony characteristics and symbiotic effectiveness of Rhizobium. J. Appl. Bact. 38:19-27.
59. Heumann,
W. 1968. Conjugation in starforming Rhizobium lupini.
Mol. Gen. Genet. 102:132-144.
60.
Higashi,
S. 1967. Transfer of clover infectivity of Rhizobium trifolii
to Rhizobium phaseoli as mediated by an episomic factor. J. Gen.
Appl. Microbiol. 13:391403.
61. Hirsch, P. R. 1979. Plasmid-determined
bacteriocin production by Rhizobium leguminosarum. J. Gen. Microbiol. 113:219-228.
62.
Hirsch,
P. R., M. Van Montagu, A. W. B. Johnston, N. J. Brewin, and J. Schell. 1980. Physical
identification of bacteriocinogenic, nodulation, and other plasmids in strains
of Rhizobium leguminosarum.
J. Gen. Microbiol. 120:403-412.
63.
Hoagland,
D. R., and D. I. Arnon. 1938.
The water culture method for growing plants without soil. California A9,
Exp. Std. Circular Number 347.
64.
Hollis,
A. B., W. E. Kloos, and G. H. Elkan. 1981. DNA: DNA hybridization studies of Rhizobium
japonicum and
65. Hombrecher, G., N. J. Brewin, and A. W. B.
Johnston. 1981. Linkage of genes for nitrogenase and
nodulation ability on plasmids in Rhizobium leguminosarum and R.
phaseoli. Mol. Gen. Genet.
182:133-136.
66. Hooykaas,
P. J. J., A. A. N. van Brussel, H. den Dulk-Ras, G. M. S. van Slogteren, and
R. A. Schilperoort. 1981. Sym plasmid of Rhizobium trifolii
expressed in different rhizobial species and Agrobacterium tumefaciens. Nature 291:351-353.
67. Hughes,
D. Q., and J. M. Vincent.
1942. Serological studies of the
root-nodule bacteria. III. Tests of neighboring strains of the same
species. Proc. Linnean Soc. N.S. Wales.
67:142-152.
68. Humphrey, B. A., and J. M. Vincent. 1962.
Calcium in cell walls of Rhizobium trifolii. J. Gen. Microbiol. 29:557-561.
69. Humphrey,
B. A., and J. M. Vincent.
1965. The effect of calcium
nutrition on the production of diffusible antigens by Rhizobium trifolii. J. Gen. Microbiol. 41:109-118.
70. Humphrey, B. A., and J. M. Vincent. 1969.
The somatic antigens of two strains Rhizobium trifolii. J. Gen. Microbiol. 59:411-425.
71. Humphrey, B. A., and J. M. Vincent. 1975.
Specific and shared antigens in strains of Rhizobium meliloti. Microbios. 13:71-76.
72. Hunter, C. A., and H. G. Crecelius. 1938.
Detection of hydrogen sulphide in cultures. J. Bacteriol. 35:185-196.
73. Jarvis, B. D. W., C. E. Pankhurst, and J. J.
Patel. 1982. Rhizobium loti a new species
of legume root nodule bacteria. Int. J.
Syst. Bacteriol. 32:378-380.
74. Johnson, G. V., H. J. Evans, and T. Ching. 1966.
Enzymes of the glyoxylate cycle in rhizobia and nodules of legumes. Pl. Physiol. 41:1330-1336.
75. Johnson, K. M., and J. McN. Sieburth. 1977.
Dissolved carbohydrates in seawater:I, a precise spectrophotometric
analysis for monosaccharides. Marine
Chem. 5:1-13.
76. Johnston, A. W. B., J. L. Beynon, A. V.
Buchanan-Wollaston, S. M. Setchell, P. R. Hirsch., and J. E. Beringer. 1978.
High frequency transfer of nodulation ability between strains and
species of Rhizobium. Nature
276:634-636.
77. Johnston, A. W. B., G. Hombrecher, A. J.
Brewin, and M. C. Cooper.
1982. Two transmissible plasmids
in Rhizobium leguminosarum strain 300. J. Gen. Microbiol.
128:85-93
78. Jones,
D. G., and P. E. Russel. 1972. The application of immunofluorescence
techniques to host plant/nodule bacteria selectivity experiments using Trifolium
repens. Soil Biol. Bioehem.
4:277-282.
79. Jordan,
D. C. 1982. Transfer of Rhizobium japonicum
Buchanan 1980 to Bradyrhizobium gen. nov., a genus of slow-growing root
nodule bacteria from leguminous plants.
Int. J. Syst. Bacteriol. 32:136-139.
80. Jordan,
D. C., and O. N. Allen. 1974. Genus II. Rhizobium, p. 262-264. In R. E. Buchanan and N. E. Gibbons
(ed.), Bergey’s Manual of Determinative Bacteriology, 8th ed. The Williams and Wilkins Co., Baltimore.
81. Josey,
D. P., J. L. Beynon, A. W. B. Johnston, and J. E. Beringer. 1979.
Strain identification in Rhizobium using intrinsic antibiotic
resistance. J. Appl. Bacteriol.
46:343-350.
82. Keele,
B. B. Jr., P. B. Hamilton, and G. H. Elkan. 1969. Glucose catabolism
in Rhizobium japonicum.
J. Bacteriol. 97:1184-1191.
83. Kennedy,
C., B. Dreyfus, and J. Brockwell.
1981. Transfer, maintenance and
expression of P plasmids in strains of cowpea rhizobia. J. Gen. Microbiol. 125:233-240.
84. Keyser,
H. H., B. B. Bohlool, T. S. Hu, and D. F. Weber. 1982. Fast-grwoing
rhizobia isolated from root nodules of soybean. Science 215:1631-1632.
85. Kingsley,
M. T., and B. B. Bohlool.
1981. Release of Rhizobium spp.
from tropical soils and recovery for immunofluorescence enumeration. Appl. Environ. Microbiol. 42:241-248.
86. Kleczkowski,
A., and H. G. Thorton. 1944. A serological study root-nodule bacteria
from pea and clover inoculation groups.
J. Bacteriol. 48:661-672.
87. Kondorosi,
A., G. B. Kliss, T. Forrai, B. Vincze, and Z. Banfalvi. 1977.
Circular linkage map of the Rhizobium meliloti chromosome. Nature 268:525-527.
88. Koontz,
F. P., and J. E. Faber. 1961. Somatic antigens of Rhizobium japonicum. Soil Sci. 91:228-232.
89. Koser,
S. A. 1923. Utilisation of the salts of organic acids by
the colon-aerogenes group. J.
Bacteriol. 8:493
90. Kovaks,
N. 1956. Identification of Pseudomonas pyocyanea by the
oxidase reaction. Nature 128:703.
91. Kowalski, M. 1967. Transduction in Rhizobium
meliloti. Acta Microbiol. Polon.
16:7-12.
92. Krasilnikov, N. A. 1941.
Variability of nodule bacteria. Doklady Akademii Nauk SSSR
31:90-92. (In Russian).
93. Krasilnikov, N. A. 1945.
Induction of new virulence properties in nodule bacteria. Mikrobiol.
14:230-236. (In Russian).
94. Krol, A. J. N., J. G. G. Hontelez, R. C.
Van Den Bos, and A. Van Kamen.
1981. Expression of large
plasmids in the endosymbiotic form of Rhizobium leguminosarum. Nucleic Acid Res. 8:4337-4347.
95. Kuykendall, L. D. 1979.
Transfer of R factors to and between genetically marked sublines of Rhizobium
japonicum. Appl. Environ.
Microbiol. 37:862-866.
96. Leonard, L. T. 1943.
A simple assembly for use in the testing of cultures of rhizobia. J. Bacteriol. 45:523-527.
97. Lohnis, F., and R. Hansen. 1921.
Nodule bacteria of leguminous plants.
J. Agr. Res. 20:543-555.
98. Lowry, 0. H., N. J. Rosenbrough, A. L.
Farr, and R. J. Randall. 1951. Protein measurement with the Folin phenol
reagent. J. Biol. Chem. 193:265-275.
99. Maier,
R. J., and W. J. Brill. 1976. Ineffective and non-nodulatiog mutant
strains of Rhizobium japonicum.
J. Bacteriol. 127:763-769.
100. Martinez-de
Drets, G., and A. Arias. 1970. Metabolism of some polyols by Rhizobium
meliloti. J. Bacteriol.
103:97-103.
101. Martinez-de
Drets, G., and A. Arias. 1972. Enzymatic basis for the differentiation of Rhizobium
into fast- and slow-growing groups. J.
Bacteriol. 109:467-470.
102. Martinez-de
Drets, G., and A. Arias. 1977. 6-Phospho-D-gluconate:
NAD+ 2-oxidoreductase (decarboxylating) from slow-growing
rhizobia. J. Bacteriol. 130:1139-1143.
103. Masterson,
R. V., P. R. Russell, and A. G. Atherly.
1982. Nitrogen fixation (nif)
genes and large plasmids of Rhizobium japonicum. J. Bacteriol. 152:928-931.
104. May,
S. N., and B. B. Bohlool.
1983. Competition among Rhizobium
leguminosarum strains for nodulation of lentils (Lens esculenta).
Appl. Environ. Microbiol. 45:960-965.
105. Morrison,
N. A., Y. H. Cen, M. J. Trinick, J. Shine, and B. G. Rolfe. 1983.
Heat curing of a Sym plasmid in a fast-growing Rhizobium sp. that
is able to nodulate legumes and the nonlegume Parasponia sp. J. Bacteriol. 153:527-531.
106. Norris,
D. 0. 1965. Acid production by Rhizobium: an
unifying concept. Pl. Soil 22:143-166.
107. Nuti, M., A. M. Ledeboer, A. A. Lepidi, and
R. A. Schilperoort. 1977. Large plasmids in different Rhizobium species. J. Gen. Microbiol. 100:241-248.
108. Nuti,
M. P., A. A. Lepidi, R. K. Prakash, R. A. Schilperoort, and F. C. Cannon. 1979.
Evidence for nitrogen fixation (nif) genes on indigenous Rhizobium
plasmids. Nature 282:533-535.
109. Page, W. J. 1978. Transformation of Azotobacter vinelandii
strains unable to fix nitrogen with Rhizobium spp. DNA. Can J. Microbiol. 24:209-214.
110. Pankhurst,
C. E. 1979. Some antigenic properties of cultured cells
and bacteroid forms of fast- and slow-growing strains of Lotus
rhizobia. Microbios. 24:19-28.
111. Pariiskaya,
A. N. 1973. The effect of acridine orange and mitomycin
C on the symbiotic properties of Rhizobium meliloti. Mikrobiol.
42:119-121.
112. Peterson, J. B., and T. LaRue. 1981. Utilization of aldehydes and alcohols by soybean bacteroids. Pl. Physiol. 68:489-493.
113. Pilicinski, W. P., and E. L. Schmidt. 1981.
Plasmid transfer within and between serologieally distinct strains of Rhizobium
japonicum, using antibiotic resistance mutants and auxotrophs. J. Bacteriol. 145:1025-1030.
114. Prakash, R. K., P. J. J. Hooykaas, A. M.
Ledeboer, J. W. Kijne, R. A. Schilperoort, M. P. Nuti, A. A. Lepidi, F. Casse,
C. Boucher, J. S. Julliot, and J. Denarie.
1980.
Detection, isolation, and characterization of large plasmids in Rhizobium,
p. 139-163.
In W. E. Newton and W. H. Orme-Johnson (ed.), Nitrogen
Fixation. University Park Press, Baltimore.
115.
Raina,
J. L., and V. V. Modi. 1969.
Genetic transformation in Rhizobium. J. Gen. Microbiol. 57:125-130.
116. Raina, J. L., and V. V. Modi. 1971.
Further studies on genetic transformation in Rhizobium. J. Gen. Microbiol. 65:161-165.
117.
Raina,
J. L., and V. V. Modi. 1972.
Deoxyribonucleate binding and transformation in Rhizobium japonicum. J. Bacteriol. 111:356-360.
118. Rigaud, J., and J. Trinchant. 1973. Isolement d’une alcool deshydrogenase
chez Rhizobium et role dans le metabolisme indolique. Physiol. Plant. 23:160-165.
119. Roberts,
G. P., W. T. Leps, L. E. Silver, and W. J. Brill. 1980. Use of
two-dimensional polyacrylamide gel electrophoresis to identify and classify Rhizobium
strains. Appl. Environ. Microbiol. 39:414-422.
120. Ruvkin,
G. B., and F. M. Ausubel. 1980.
Interspecies homology of nitrogenase genes. Proc. Nat. Acad. Sci. U.S.A. 77:191-195.
121. Schmidt,
E. L. 1973. Fluorescent antibody techniques for
the study of microbial ecology. Bull.
Eco. Res. Comm. (Stockholm). 19:67-76.
122. Schmidt,
E. L. 1974. Quantitative autecological study of
microorganisms in soil by immunofluorescence.
Soil Sci. 118:141-149.
123. Schmidt,
E. L., R. 0. Bankole, and B. B. Bohlool. 1968. Fluorescent antibody approach to the study
rhizobia in soil. J. Bacteriol. 95:1987-1992.
124. Skrdleta,
V. 1965. Somatic sero-groups of Rhizobium
japonicum. Plant Soil 23:43-48.
125. Skrdleta,
V. 1969. Serological analysis of eleven
strains of Rhizobium japonicum.
Antonie. van Leeuwenhoek 35:77-83.
126. Smibert,
R. M., and N. R. Krieg. 1981.
General characterizations, p. 409-443. In P. Gerhardt (chief ed.),
Manual of Methods for General Bacteriology.
American Society for Microbiology, Washington, D. C.
127. Stevens,
J. W. 1923. Can all strains of a specific
organism be recognized by agglutination?
J. Infectious Diseases. 33:557-566.
128. Strzelcowa,
A. 1968. The use of the technique of van
Schreven for the taxonomy of Rhizobium strains. Acta Microbiol. Polon. 17:263-268.
129. Tajima,
S., and T. A. LaRue. 1982.
Enzymes for acetaldehyde and ethanol formation in legume nodules. Plant Physiol. 70:388-392.
130. Trinick,
M. J. 1965. Medicago sativa
nodulation with Leucaena leucocephala root nodule bacteria. Aust. J. Sci.
27:263-264.
131. Van
Straten, J., and E. L. Schmidt. 1974.
Volatile compounds produced during acetylene reduction by detached
soybean nodules. Soil Biol. Biochem. 6:347-351.
132.
Vincent, J. M. 1970. A Manual for the Practical Study of
Root-Nodule Bacteria. IBP Handbook 15. Blackwell
Scientific Publications, Oxford.
133. Vincent,
J. M. 1974.
Root-nodule symbiosis
with Rhizobium, p. 266-341. In A. Quispel (ed.), Biology
of Nitrogen Fixation. North Holland Publishing Co., Amsterdam.
134. Vincent,
J. M. 1977.
Rhizobium: General microbiology, p. 277-366. In
R. W. F. Hardy and W. S. Silver (ed.), A Treatise on Dinitrogen
Fixation, Sec. III Biology. John Wiley
and Sons, New York.
135. Vincent,
J. M. 1982.
The basic serology of
rhizobia, p. 13-26. In J. M. Vincent (ed.), Nitrogen Fixation in
Legumes. Academic Press, Sydney.
136. Vincent,
J. M., and B. Humphrey. 1970. Taxonomically significant group
antigens in Rhizobium. J. Gen.
Microbiol. 63:379-382.
137. Vincent,
J. M., B. Humphrey, and V. Skrdleta. 1973. Group antigens in slow-growing
rhizobia. Arch. Mikrobiol. 89:79-82.
138. Vincent,
J. M., P. S. Nutman, and F. A. Skinner. 1979. The identification and classification of Rhizobium,
p. 49-69.
In F. A. Skinner and D. W. Lovelock (ed.), Identification
Methods for Microbiology, 2nd ed.
Academic Press, London.
139. Vintikova,
H., M. Srogl, and V. Skrdleta. 1961. A contribution
to the serological typization of the rhizobia.
Folia Microbiol. 6:243-249.
140. Wright, W. H. 1925. The nodule bacteria of
soybeans: I. Bacteriology of
strains. Soil Sci. 20:95-141.
141. Zurkowski, W., M. Hoffman, and Z. Lorkiewicz. 1973. Effect of
acriflavin and sodium dodecyl sulphate on infectiveness of Rhizobium trifolii. Acta Microbiol. Polon. 5:55-60.
142. Zurkowski, W., and Z. Lorkiewicz. 1978. Effective
method for the isolation of non-nodulatiog mutants of Rhizobium trifolii.
Genet. Res. 32:311-314.
143. Zurkowski, Y., and Z.
Lorkiewicz. 1979.
Plasmid-mediated control of nodulation in Rhizobium trifolii. Arch. Microbiol. 123:195-201.