







Impact of rising temperature on carbon input, partitioning, loss and storage in tropical wet forests

Figure 1a. One of nine permanent plots in tropical wet forest on the Island of Hawaii.
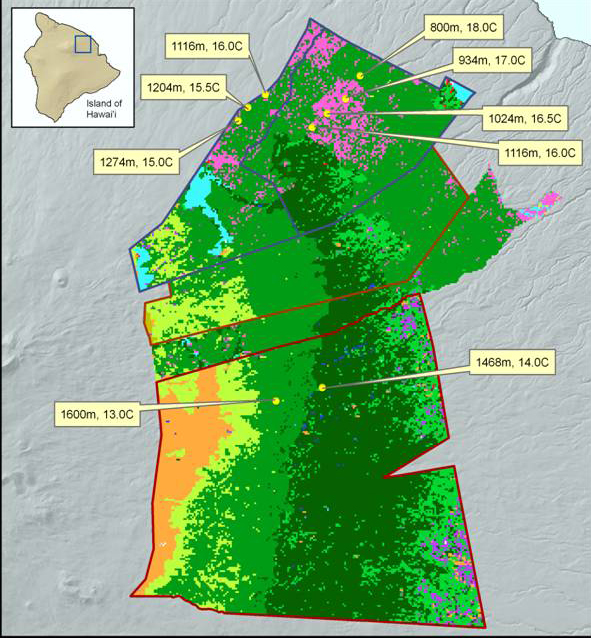
Figure 1b. Location of nine permanent plots across a 5.2°C mean annual temperature gradient. Plots are located in The Hawaii Experimental Tropical Forest (Laupahoehoe Unit) of the USDA Forest Service, and the Hakalau
Forest National Wildlife Refugee of the US Fish and Wildlife Service.
In collaboration with Dr. Christian Giardina of the Institute of
Pacific Islands Forestry, we are conducting a
study to examine
how rising temperatures will impact carbon cycling in
tropical wet forest
ecosystems. Terrestrial ecosystem carbon storage in soils and
vegetation exceeds that in the atmosphere by a factor of four, and
represents a dynamic balance among carbon input,
partitioning, loss, and storage. This balance is likely
being altered by climate change, but
the response of terrestrial carbon cycling to rising temperatures
remains poorly quantified. Importantly, temperature-induced
changes in ecosystem carbon flux and storage have the potential to
feedback into atmospheric CO2
levels and global climate. In this study we are examining how
tropical forest ecosystems will respond to rising temperature by
examining ecosystem carbon storage (live biomass, coarse woody debris
and soil organic matter); carbon input (gross primary production; GPP);
carbon fluxes (litterfall, aboveground net primary productivity (ANPP),
soil-surface CO2 efflux, and total belowground carbon flux
(TBCF)); and carbon partitioning (fraction of GPP partitioned to
aboveground vs. belowground) across a 5.2°C mean annual temperature
(MAT) gradient on the Island of Hawaii. Along the MAT gradient,
substrate type and age, dominant overstory vegetation, disturbance history, and plant
available water are constant, allowing us to isolate the impacts of
temperature on ecosystem carbon cycling. This work is funded by the National Science Foundation, the USDA Forest
Service, and the College of Tropical Agriculture and Human Resources, University of Hawaii at Manoa.
If you are interested in conducting research utilizing our existing experimental design across the MAT gradient, please complete and email this form to litton@hawaii.edu.
Publications to date
Bothwell L, Selmants, PC, Giardina, CP, Litton, CM
(In review) Leaf litter decomposition rates increase with rising mean
annual temperature in Hawaiian tropical montane wet forests. PeerJ.
Giardina CP, Litton CM,
Crow SE, Asner GP (2014) Increased total belowground carbon flux,
and not soil carbon loss, drives temperature related increases in soil
respiration. Nature Climate Change 4: 822-827. (PDF)
Selmants PC, Litton CM,
Giardina CP, Asner, GP (2014) Ecosystem carbon storage does not
vary with mean annual temperature in Hawaiian tropical montane wet
forests. Global Change Biology 20: 2927-2937. (PDF)
Mascaro J, Litton CM,
Hughes RF, Uowolo A, Schnitzer SA (2014) Is logarithmic transformation
necessary in allometry? Ten, one-hundred, one-thousand-times yes.
Biological Journal of the Linnean Society, 111, 230-233. (PDF)
Iwashita DK, Litton CM,
Giardina CP (2013) Coarse woody debris carbon storage across a mean
annual temperature gradient in tropical montane wet forest. Forest
Ecology and Management, 291, 336-343. (PDF)
Litton CM,
Giardina CP, Albano JK, Long MS, Asner GP (2011) The magnitude and
variability of soil-surface CO2 efflux increase with temperature in
Hawaiian tropical montane wet forests. Soil Biology & Biochemistry,
43, 2315-2323. (PDF)
Mascaro J, Litton CM,
Hughes RF, Uowolo A, Schnitzer SA (2011) Minimizing bias in biomass
allometry: Model selection and log-transformation of data. Biotropica,
43, 649-653. (PDF)
Ise T, Litton CM, Giardina CP,
Ito A (2010) Comparison of modeling approaches for carbon partitioning:
Impact on estimates of global net primary production and equilibrium
biomass of woody vegetation from MODIS GPP. Journal of Geophysical Research-Biogeosciences, 115, G04025, doi:10.1029/2010JG001326. (PDF)
Litton CM, Giardina
CP (2008) Belowground carbon flux and partitioning: Global patterns and
response to temperature (Invited article). Functional Ecology, 22, 941-954. (PDF)
Global patterns in carbon flux and partitioning
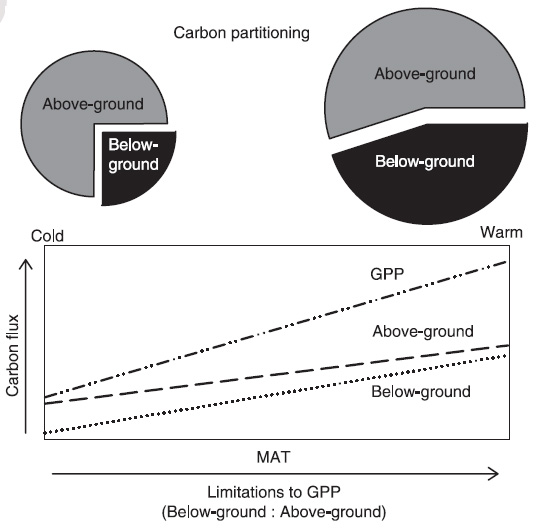
Figure 2. Hypothesized relationship between mean annual
temperature and the partitioning of GPP (carbon flux as a fraction of
GPP) to aboveground vs. belowground (top panel). While GPP,
aboveground C flux, and belowground C flux all increase with MAT, the
slopes of the aboveground and belowground relationships differ
because the factors constraining GPP change as MAT increases (bottom
panel). At colder sites, air temperature presents the strongest
limitation to GPP, and belowground resource supply (e.g.,
nutrients and water) is high by comparison. Conversely, at warmer
sites, air temperature constraints are alleviated and belowground
resource supply exerts a stronger limitation to GPP. As a result,
partitioning of GPP to belowground increases at higher MAT (from
Litton and Giardina 2008).
Carbon allocation plays a critical role in forest ecosystem carbon
cycling by shifting the products of photosynthesis between respiration
and biomass production, ephemeral and long-lived tissues, and
aboveground and belowground components. As a primary control on
terrestrial carbon storage and forest ecosystem carbon dynamics, carbon
allocation is a dynamic balance among total ecosystem carbon input
(gross primary production), carbon fluxes and partitioning of GPP
to individual components, and carbon loss. Some of our recent work on carbon
allocation has focused on global syntheses of available data in forest ecosystems. This work is designed to inform terrestrial ecosystem models by examining general patterns in carbon flux and partitioning, and their
response to variables such as resource availability, stand age, competition, and climate change.
Publications to date
Ise T, Litton CM, Giardina CP,
Ito A (2010) Comparison of modeling approaches for carbon partitioning:
Impact on estimates of global net primary production and equilibrium
biomass of woody vegetation from MODIS GPP. Journal of Geophysical Research-Biogeosciences, 115, G04025, doi:10.1029/2010JG001326. (PDF)
Litton CM, Giardina
CP (2008) Belowground carbon flux and partitioning: Global patterns and
response to temperature (Invited article). Functional Ecology 22: 941-954. (PDF)
Litton CM, Raich JW, Ryan MG (2007) Review: Carbon allocation in forest ecosystems. Global Change Biology, 13, 2089-2109. (PDF)
Invasive grasses, wildfire, and native forest restoration on Oahu

Figure 3a. The spread of
invasive species, particularly nonnative grasses, and repeated
wildfires, both accidental and prescribed, have converted much of
Hawaii's dry forest ecosystems to nonnative grasslands.

Figure 3b. Guinea
grass (Urochloa maxima), a nonnative invasive grass in Hawaii, forms dense
stands that outcompete native plants and has very high fine fuel loads that greatly increase fire
potential, spread, and severity.

Figure 3c. A restoration trial with native species assemblages on a guinea grass dominated site, designed to
simultaneously restore native biodiversity and decrease
the probability and severity of future fire.
Wildland
fires are a significant problem in Hawaiian landscapes where native
woody communities have been replaced by nonnative invasive
grasses. Wildfires burning in areas dominated by invasive
grasses, such as guinea grass (Megathyrsus maximus),
typically degrade remnant native plant communites, and preclude the
estabilshment and restoration of native species assemblages. Many
wildfires on military lands are ignited by training activities, with
subsequent negative impacts on training opportunities and military
preparedness. In order to simultaneously train on military lands
and protect remnant native species and communities, the invasive
grass-wildfire cycle needs to be managed and ultimately eliminated. Our
work is designed to provide a better understanding of the fuel,
climatic, and fire behavior components of the invasive grass-wildfire
cycle in Hawaiian dry ecosystems currently dominated by nonnative
grasses. Specifically, this work is designed to improve models to
accurately predict the probability of ignition, rate of spread, and
fire intensity in guinea grasslands. In addition, we are exploring methods to restore
native woody plant communities to these areas to reduce the likelihood
of fire occurrence and spread, to eliminate further conversion of
remnant native plant communities to nonnative grasslands, and to
increase native biodiversity in these highly degraded ecosystems. This work is
funded by the Department of Defense (U.S. Army Garrison - Oahu), the
USDA Forest Service (National Fire Plan), and the the College of Tropical Agriculture and Human Resources - University of Hawaii at Manoa.
Publications to date
Pierce A, McDaniel S, Wasser M, Ainsworth A, Litton CM,
Giardina CP, Cordell S (In press) Using a free-burning prescribed
fire to test custom and standard fuel models for fire behavior
prediction in a grass-invaded tropical dry shrubland. Applied
Vegetation Science: doi:10.1111/avsc.12111.
Ellsworth LM, Litton CM,
Dale A, Miura T (In press) The grass-fire cycle at a landscape
scale: Changes in land cover and fire behavior with nonnative grass
invasion on a tropical island. Applied Vegetation Science:
doi:10.1111/avsc.12110s.
Ammondt SA, Litton CM,
Ellsworth LM, Leary JK (2013) Restoration of native plant communities
in a Hawaiian dry lowland ecosystem dominated by the invasive grass Megathyrsus maximus. Applied Vegetation Science, 16, 29-39. (PDF)
Ellsworth LM, Litton CM, Taylor AD, Kauffman JB (2013) Spatial and temporal variability of guinea grass (Megathyrsus maximus) fuel loads and moisture on Oahu, Hawaii. International Journal of Wildland Fire, 22, 1083-1092. (PDF)
Ammondt SA, Litton CM
(2012) Competition between native Hawaiian plants and the invasive
grass Megathyrsus maximus: Implications of functional diversity for
ecological restoration. Restoration Ecology, 20, 638-646. (PDF)
Impacts of nonnative ungulates on ecosystem structure and function in Hawaiian forests

Figure 4a. Nonnative feral pigs degrade native wet forests in Hawai‘i via rooting and mixing
of soil horizons, trampling and
consumption of native plants, and transport of nonnative seeds (Photo Credit: Hawaii Volcanoes National Park).

Figure 4b. Feral pig damage in native wet forests on the Island of Hawai‘i. This fence-line photo, taken only six months after the construction of a pig-proof fence, demonstrates
pig disturbance to the soil surface. The left-side of the image (where
pigs are still present) has almost no litter layer and limited plant
establishment, while the right-side of the image (where pigs have been
excluded) has an intact litter layer and abundant native forest
regeneration.
Hawaiian forest ecosystems are currently undergoing rapid degradation as a result of nonnative feral ungulates,
with implications for forest composition, structure, and
biogeochemistry. For example, feral pigs disturb soil via rooting and mixing of soil
horizons, and plant communities via the trampling and consumption of
native plants and transport of nonnative seeds. We are
examining how dominant nonnative ungulates, and their subsequent
removal, alters ecosystem structure and composition in native Hawaiian wet and dry forest ecosystems. Our
current work is focused on: (1) understanding native and nonnative plant
community dynamics in the presence of nonnative ungulates, and following their
removal; (2) examining
the biogeochemical impacts
of nonnative ungulates in these forests, and the response to their
removal; and (3) testing management strategies to improve native
vegetation following nonnative ungulate removal. This work hass been
funded by the
USDA-CSREES-TSTAR Pacific Program, the DoD Strategic Environmental
Research and Development Program (SERDP), and the College of Tropical Agriculture and Human Resources, University of Hawaii at Manoa.
Publications to date
Cole RJ, Litton CM (2014)
Vegetation response to removal of non-native feral pigs from Hawaiian
tropical montane wet forest. Biological Invasions, 16, 125-140. (PDF)
Murphy
MJ, Inman-Narahari F, Ostertag R, Litton CM (2014) Invasive feral pigs
impact native tree ferns and woody seedlings in Hawaiian forest.
Biological Invasions, 16, 63-71. (PDF)
Chynoweth MW, Litton CM, Lepczyk CA, Hess SC, Cordell S (2013) Biology and impacts of Pacific Island invasive species. 9. Capra hircus, the feral goat (Mammalia: Bovidae). Pacific Science 67: 141-156. (PDF)
Cole RJ, Litton CM,
Koontz MJ, Loh RK (2012) Vegetation recovery 16 years after feral pig
removal from a wet Hawaiian forest. Biotropica, 44, 463-471. (PDF)
Dunkell DO, Bruland GL,
Evensen CI, Litton CM (2011) Runoff, sediment transport, and effects of
feral pig (Sus scrofa) exclusion in a forested Hawaiian watershed.
Pacific Science, 65, 175-194. (PDF)
Ecosystem-level impacts of nonnative grass invasion in Hawaiian Dry Forests

Figure 5. We are
using invasion of Hawaiian dry forests by a nonnative perennial
bunchgrass to understand the impacts of invasion on carbon
cycling, water availability and use, and nutrient dynamics.
The image at the left shows one of our plots where the nonnative
grass has been removed from the understory.
Nonnative invasive plants are prevalent in terrestrial ecosystems
worldwide and have long been recognized to adversely impact native
species assemblages and biodiversity. However, it is only in the
past several decades that invasions have been implicated as driving
changes in important ecosystem processes. During this time
progressively more attention has been focused on the consequences of
invasions for ecosystem function, with particular emphasis on soil
nutrient cycling. We are using invasion of Hawaiian dry forest by
an African perennial bunchgrass (Pennisetum setaceum) to examine how nonnative invasion into forest ecosystems impacts: (i) water availability and use; (ii) aboveground and belowground ecosystem carbon pools, fluxes and partitioning; and (iii)
nutrient dynamics. The majority of this work is being conducted at the Kaupulehu Dry Forest Preserve, in a
series of canopy-intact plots established in 2000, where the invasive
grass understory was removed from half of the plots and left intact in
the remaining half.
Publications to date
Thaxton JM, Cole TC, Cordell S, Cabin RJ, Sandquist DR, Litton CM
(2010) Native species regeneration following ungulate exclusion and
nonnative grass removal in a remnant Hawaiian dry forest. Pacific
Science, 64, 533-544. (PDF)
Litton CM, Sandquist
DR, Cordell S (2008) A nonnative invasive grass increases soil carbon
flux in a Hawaiian tropical dry forest. Global Change Biology, 14 726-739. (PDF)
Litton CM, Sandquist DR, Cordell S (2006) Effects of non-native grass
invasion on aboveground carbon pools and tree population structure in a
tropical dry forest of Hawaii. Forest Ecology and Management, 231, 105-113. (PDF)
Impact of fire, invasive species, and their interactions on carbon cycling in tropical rainforests

Figure 6.
We are using a natural elevation/precipitation gradient in Hawaii
Volcanoes National Park to examine how lava-ignited
wildfires, nonnative invasive species, and their
interactions impact: (i) aboveground carbon sequestration in
vegetation and detritus, and (ii) fuel loads and fire behaviour.
Fire is increasingly recognized as an important natural disturbance in
the tropics. However, little is known about the evolutionary
history of fire in shaping the structure and function of tropical
rainforests. In addition, many tropical forests are now heavily
impacted by nonnative species which can disrupt ecosystem processes and services, and alter successional trajectories
and disturbance regimes.
We are examining the synergistic impacts of lava-ignited wildfire and
nonnative species invasions on aboveground carbon pools in vegetation
and detritus along a precipitation gradient in Hawaii Volcanoes
National Park. Potential changes in carbon sequestration in
tropical forests as a result of wildfire and nonnative species
interactions are particularly important in light of the ubiquitous
presence of invasive species and the need for better understanding of
the role they will play in disturbance regimes and global C cycling.
Future work will concentrate on understanding how invasive
species impact fuel loading and fire behaviour in this system.
Publications to date
Litton CM, Kauffman JB. Impact of fire, invasive species, and their interactions on aboveground carbon cycling in tropical mesic to wet rainforests. Forest Ecology and Management, In prep.
Ainsworth A, Kauffman JB
(2009) Response of native Hawaiian woody species to lava-ignited
wildfires in tropical forests and shrublands. Plant Ecology, 201 197-209.
Litton CM, Kauffman JB (2008)
Allometric models for predicting aboveground biomass in two widespread
woody plants in Hawaii, U.S.A. Biotropica, 40 313-320. (PDF)
Impact of fire on plant community dynamics, soils, and ecosystem processes in native forests of south-central Chile

Figure 7.
We are working in endemic Nothofagus glauca forests in south-central Chile to examine the impacts of wildfire, and subsequent invasion by the nonnative Pinus radiata, on plant community dynamics and ecosystem processes.
The temperate deciduous species Nothofagus glauca, endemic to Chile, exhibits characteristics commonly
found in fire-adapted vegetation, yet the role of fire in the
evolutionary history of the vegetation in this area is poorly understood. We are examining the effects of wildfire
on secondary postfire succession in a N. glauca
forest in the Coastal Cordillera of south-central Chile. Our work has documented that the
majority of the plants associated with this forest type exhibit
adaptations to survive fire and/or colonize the postfire environment. However, the presence and success of
exotic invaders, particularly Pinus radiata,
is altering the successional trajectory of this endemic community with unknown implications for important ecosystem processes.
Currently we are studying how invasion of these forests by P. radiata is impacting water availability and use.
Publications to date
Litton CM, Santelices R, Sandquist DR. Pinus radiata invasion following fire alters water availability in Nothofagus glauca forests of south-central Chile. Plant Ecology, In prep.
Litton CM, Santelices R (2003) Effect of wildfire on soil physical and chemical properties in a Nothofagus glauca forest, Chile. Revista Chilena de Historia Natural, 76, 529-542. (PDF)
Litton CM, Santelices R (2002) Early post-fire succession in a Nothofagus glauca forest in the Costal Cordillera of south-central Chile. International Journal of Wildland Fire, 11, 115-125. (PDF)
Impact of fire, as a natural disturance, on carbon cycling in lodgepole pine forests

Figure 8.
Fire is a natural disturbance in most forest ecosystems that drives tremendous spatial heterogeneity across landscapes.
We are examining how fire impacts carbon pools and fluxes
across Rocky Mountain landscapes through postfire legacies
in stand age and tree density.
Validating the different components of the carbon budget in forest
ecosystems is essential for developing allocation rules that allow
accurate predictions of global carbon pools and fluxes. In
addition, a better understanding of the effects of natural disturbances
on carbon cycling is critical – particularly in light of changes
in disturbance regimes that may occur with alterations in global
climate. This study investigated the indirect effects of fire on
carbon cycling in lodgepole pine (Pinus contorta var. latifolia
Engelm. ex Wats.) stands in Yellowstone National Park by examining
aboveground and belowground carbon pools, fluxes and allocation
patterns in post-fire stands that varied in tree density and stand age
(four forest types: low (<1000 trees/ha), moderate
(7,000–40,000 trees/ha), and high tree densities (>50,000
trees/ha) in 13-yr-old stands; and ~110-yr-old mature stands).
Publications to date
Litton CM, Ryan MG, Knight DH (2004)
Effects of tree density and stand age on carbon allocation patterns in
postfire lodgepole pine. Ecological Applications, 14, 460-475. (PDF)
Turner MG, Tinker DB, Romme WH, Kashian DM, Litton CM (2004) Landscape
patterns of sapling density, leaf area, and aboveground net primary
production in postfire lodgepole pine forests, Yellowstone National
Park (USA). Ecosystems, 7, 751-775. (PDF)
Litton CM, Ryan MG, Knight DH, Stahl PD (2003) Soil-surface CO2 efflux
and microbial biomass in relation to tree density thirteen years after
a stand replacing fire in a lodgepole pine ecosystem. Global Change Biology, 9, 680-696. (PDF)
Litton CM, Ryan MG, Tinker DB, Knight DH (2003) Belowground and
aboveground biomass in young postfire lodgepole pine forests of
contrasting tree density. Canadian Journal of Forest Research, 33, 351-363. (PDF)